EKSPRESI GEN Gα dan PEROKSIDASE PADA KEDELAI KULTIVAR LUMUT YANG MENDAPAT CEKAMAN ALUMINIUM RIZKI AMELIA LUBIS A351030211 SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2008

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

EKSPRESI GEN Gα dan PEROKSIDASE PADA KEDELAI
KULTIVAR LUMUT YANG MENDAPAT CEKAMAN
ALUMINIUM
RIZKI AMELIA LUBIS
A351030211
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2008

SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul :
Ekspresi Gen Heterotrimerik Gα dan Peroksidase pada Tanaman Kedelai
Kultivar Lumut Yang Mendapat Cekaman Aluminium
Adalah benar merupakan hasil karya saya sendiri yang diarahkan oleh Komisi
Pembimbing dan belum pernah dipublikasikan untuk kepentingan lain. Semua sumber
data dan informasi yang digunakan telah dinyatakan secara benar dan dapat diperiksa
kebenarannya.
Bogor, Februari 2008
Rizki Amelia Lubis A351030211

RINGKASAN
RIZKI AMELIA LUBIS. Ekspresi Gen Heterotrimerik Gα dan PER pada Kedelai Kultivar Lumut yang Mendapat Cekaman Aluminium. Dibimbing oleh DIDY SOPANDIE, SUHARSONO dan UTUT WIDYASTUTI SUHARSONO Kedelai (Glycine max (L.) Merr.) merupakan salah satu tanaman pangan yang sangat penting di Indonesia. Biji kedelai dapat digunakan sebagai sumber protein nabati dari bahan pangan dan pakan. Produksi kedelai dari tahun ke tahun cenderung meningkat, namun belum dapat mengimbangi laju peningkatan permintaan dalam negeri, sehingga impor kedelai meningkat setiap tahun. Salah satu faktor penyebab rendahnya produktivitas kedelai di Indonesia adalah terbatasnya lahan tanam di pulau Jawa. Untuk mengatasi masalah ini adalah dengan melakukan pembukaan lahan baru di pulau Jawa, sehingga akan terjadi pergeseran penggunaan lahan tanam dari lahan subur ke lahan marginal. Sebagian besar lahan marginal di Indonesia berupa lahan pasang surut, lahan salin, lahan gambut dan lahan podzolik merah kuning. Lahan Podzolik Merah Kuning memiliki pH rendah dan kandungan aluminium yang cukup tinggi dan miskin kandungan zat hara tanah seperti Mg, Ca, dan P. Aluminium diketahui sebagai salah satu faktor utama penyebab keracunan bagi tanaman yang tumbuh di tanah yang bersifat masam, dengan potensi luasan di dunia sekitar 1 × 109 hektar, mencakup daerah tropis dan subtropis (Van Wambeke 1976; Haug 1984; Moller et al. 1984). Al3+ merupakan bentuk paling toksik bagi tanaman dengan gejala umumnya adalah pertumbuhan akar terhambat, akar menjadi pendek dan menebal khususnya akar utama. Usaha peningkatan produksi kedelai pada lahan masam telah banyak dilakukan, diantaranya dengan pengapuran (Widjaja Adhi1985). Walaupun tidak dipungkiri bahwa pengapuran mampu memperbaiki kondisi tanah masam, namun dalam pelaksanaannya banyak ditemui hambatan seperti masalah distribusi kapur dan aplikasinya di lapang. Penggunaan kedelai toleran pada lahan asam dengan kelarutan aluminium tinggi merupakan salah satu alternatif dalam peningkatan produksi kedelai. Protein G merupakan salah satu protein penting yang terlibat dalam penyinalan bagi sel dalam menanggapi perubahan lingkungan. Komponen jalur ini terdiri dari reseptor terkait protein G yang berada pada permukaan membran dan protein heterotrimerik G yang mentransduksi informasi dari reseptor protein G yang teraktifasi ke efektor down stream seperti kanal ion, phospholipase C. Cekaman Al diketahui menginduksi beberapa gen yang juga berhubungan dengan sistem pertahanan terhadap pathogen seperti peroksidase dan GST. Protein G subunit α berinteraksi dengan molekul reseptor yang berlokasi di membran plasma dan terlibat di dalam banyak regulasi sinyal transduksi. Aktifitas peroksidase pada akar kedelai meningkat selama perlakuan Al. Diduga kedua gen ini terlibat dalam mekanisme toleransi tanaman terhadap cekaman aluminium. Tujuan penelitian ini adalah menguji ekspresi gen Gα dan PER kedelai kultivar Lumut yang mendapat cekaman aluminium. Kondisi cekaman dilakukan dengan menggunakan larutan hara. Cekaman yang diberikan yaitu pH 4 dan pH 4+1.6mM Al. Tanaman pada larutan hara pH 6 digunakan sebagai kontrol perlakuan. Perlakuan selama 3 × 24 jam. Ekspresi gen dianalisis pada akar tanaman pada jam ke-0, 8, 24, 48, dan 72 jam setelah perlakuan. RNA total berhasil diisolasi dari akar tanaman. Total cDNA disintesis

melalui transkriptase balik dengan menggunakan total RNA sebagai template dan oligo dT sebagai primer. Analisis ekspresi gen dilakukan pada tanaman kedelai yang mendapat perlakuan pH dan perlakuan Al secara terpisah. Perlakuan pH yang diberikan adalah pH 4 dan pH 6 sebagai kontrol. Percobaan dilakukan dengan dua ulangan. Analisis ekspresi gen Gα dan gen PER dilakukan dengan cara mengamplifikasi gen spesifik tersebut dengan menggunakan cDNA total sebagai cetakannya. Langkah dalam mencampur bahan untuk PCR gen Gα dan gen PER sama dengan PCR aktin, yang membedakan hanya primer yang disesuaikan dengan gen yang dianalisis. Kondisi PCR gen Gα sama dengan kondisi PCR aktin, sedangkan untuk kondisi PCR gen PER yaitu Pra-PCR 94°C 2 menit, denaturasi 92°C selama 30 detik, annealing/penempelan 55°C selama 30 detik, ekstensi 75°C selama 1 menit, siklus diulang sebanya 30 kali, dan pasca PCR 72°C selama 5 menit. Analisis ekspresi dilakukan dengan membandingkan intensitas cahaya pita hasil PCR gen Gα dan PER terhadap kontrol aktin dengan menggunakan perangkat lunak Digi Doc-it. Hasil penelitian ini menunjukkan bahwa ekspresi gen Gα dan Peroksidase tidak dinduksi oleh cekaman Al, namun meskipun sangat kecil ekspresi gen Peroksidase diinduksi oleh perlakuan pH 4. Kata kunci; Gα, peroksidase, aluminium, cDNA

ABSTRAK
RIZKI AMELIA LUBIS, The Expression of Gα and Peroksidase Genes in Lumut Cultivar of Soybean Under Aluminium Stress. Under direction of DIDY SOPANDIE, SUHARSONO and UTUT WIDYASTUTI SUHARSONO
Gα protein interacts with molecule reseptor located in the plasma membrane and involves in regulation of various signal transduction. Activity of Peroksidase in Glycine max roots was increased during Al treatment. We supposed that the two genes involved in the plant tolerance to Al stress. The objective of this research is to study the expression of Gα and PER genes in Lumut cultivar of soybean under aluminium stress. Stress condition was carried out in a liquid culturesystem. The plant was stressed by planting in the pH 4 and pH 4+1.6 mM. The plants cultivated in pH 6 were used as a control. The experiment was carried out for 3 X 24 hours. The expression of gene was analyzed in the primary root tips at 0, 8, 24, 48 and 72 hours after the treatment. We had successfully isolated total RNA from the root tips. Total cDNA was successfully synthesized by reverse transcription method by using total RNA as a template and oligo dT as the primer. The gene expression analysis was based on the PCR by using specific primer and total cDNA as template. This experiment showed that the expression of Gα and Peroksidase genes were not induced by Al stress, although its very small expression of Peroksidase genes was induced by pH 4. Keywords; Gα, peroksidase, aluminium, cDNA

@ Hak Cipta milik IPB, tahun 2008 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB 2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
tulis dalam bentuk apapun tanpa izin IPB

EKSPRESI GEN Gα dan PEROKSIDASE PADA KEDELAI
KULTIVAR LUMUT YANG MENDAPAT CEKAMAN
ALUMINIUM
RIZKI AMELIA LUBIS A351030211
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2008

Judul Tesis : Ekspresi Gen Gα dan Peroksidase Kedelai Kultivar Lumut yang
Mendapat Cekaman Aluminium
Nama Mahasiswa : Rizki Amelia Lubis
NIM : A351030211
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Didy Sopandie, M.Agr
Ketua Dr. Ir. Suharsono, DEA Dr. Utut W. Suharsono, MS Anggota Anggota
Diketahui Ketua Program Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, MS Prof. Dr. Ir. Khairil Anwar Notodiputro, M.Sc
Tanggal Ujian : 5 Februari 2008 Tanggal Lulus : 1 September 2008

PRAKATA
Alhamdulillah, segala puji dan syukur penulis panjatkan kepada Allah SWT
yang telah memberikan karunia, rahmat dan kemudahan sehingga penelitian dan
penulisan tesis ini dapat terselesaikan. Tesis ini ditulis berdasarkan penelitian yang
selama dua tahun ini penulis lakukan di laboratorium Biologi Molekuler dan Seluler
Tanaman, Rumah Kaca dan Laboratorium Biorin Pusat Penelitian Sumberdaya Hayati
dan Bioteknologi, IPB, Bogor.
Pada kesempatan ini penulis mengucapkan terima kasih kepada Bapak Prof.
Dr. Ir. Didy Sopandie, M.Agr, selaku ketua komisi pembimbing, Bapak Dr. Ir.
Suharsono, DEA dan Ibu Dr. Ir. Utut W. Suharsono, MS selaku anggota komisi
pembimbing, atas segala jerih payah dan waktu yang telah disediakan dalam memberi
bimbingan, serta arahan mulai dari pelaksanaan penelitian hingga penulisan hasil
penelitian ini.
Ucapan terima kasih juga penulis sampaikan kepada Rektor IPB, Direktur
Sekolah Pascasarjana IPB, Ketua Program studi Agronomi dan Bioteknologi SPs-IPB
atas kesempatan yang diberikan kepada penulis untuk mengikuti pendidikan
Pascasarjana di IPB, Bogor. Terima kasih juga ditujukan kepada proyek Hibah
Bersaing Perguruan Tinggi XII Ditjen Dikti Depdiknas a.n. Dr. Utut Widyastuti dan
Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB, atas bantuannya dalam
menyediakan biaya penelitian dan fasilitas penelitian.
Penulis mengucapkan terima kasih secara khusus untuk orang tua tercinta
ayahanda H. Ichwan Lubis (Alm) dan Ibunda Hj. Yusrita Nst, yang tanpa mengenal
lelah selalu memanjatkan doa untuk keberhasilan penulis. Kepada Keluarga H. Lahum
Lubis, SH. MM., Kakak-kakak, Abang, adikku, terima kasih atas segala perhatian,
kasih sayang, bantuan serta dorongan semangat yang diberikan kepada penulis selama
ini.
Kepada rekan-rekan sesama mahasiswa yang bekerja di laboratorium BMST
dan Biorin di PPSHB, IPB : Muti, Nana, Mashuda, Firdaus, Agustina, Wiwid, Zendi,
Dyah, Budi, Joel, Bahrelfi, Ade, Yasir, mba Yusufi, Ela, Rina, Ibu Srilis, Pak Hadi,
Ibu Ayi, Ratna, Amir, Uzi, Lulut, Popi, Amay dan Tuti, laboran di Laboratorium, Pak
Mulya, Mba Pepi dan Pak Adi, Ivo juga teman-teman Sakura 11, keluarga H. Memet

Hakim, Bapak R .Atok H, Bapak M. Iqbal Z, Team Business Development serta
rekan-rekan PT. Bakrie Sumatera Plantations, Tbk dan yang lain-lain yang tidak dapat
penulis sebutkan satu persatu, penulis mengucapkan terima kasih yang sebesar-
besarnya atas pengertian, dukungan serta kerjasamanya selama ini.
Akhirnya, penulis berharap semoga tulisan ini bermanfaat dan dapat
memberikan sumbangan bagi perkembangan pertanian kedelai di Indonesia
Bogor, Februari 2008
Rizki Amelia Lubis

RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 28 Maret 1978 dari ayah H. Ichwan
Lubis (Alm) dan Hj. Yusrita Nasution. Penulis merupakan putri kelima dari enam
bersaudara.
Penulis menyelesaikan pendidikan lanjutan atas di SMA Negeri I Medan pada
tahun 1997. Pada tahun yang sama lulus UMPTN di Fakultas Pertanian USU Program
Studi Agronomi lulus pada tahun 2002. Pada tahun 2003, penulis melanjutkan studi
Agronomi pada Sekolah Pascasarjana Institut Pertanian Bogor (IPB). Saat ini penulis
bekerja di PT. Bakrie Sumatera Plantation Tbk.

DAFTAR ISI
. Halaman DAFTAR TABEL............................................................................................ ii DAFTAR GAMBAR ....................................................................................... iii PENDAHULUAN Latar Belakang ..................................................................................... 1 Tujuan Penelitian................................................................................ 4
Hipotesis .............................................................................. 4 TINJAUAN PUSTAKA
Toksisitas dan Toleransi Aluminium pada Tanaman ........................ 5 Toksisitas Al ........................................................................ 5 Toleransi Al ......................................................................... 6
Protein heterotrimerik Gα .................................................................. 7 Gen yang berhubungan dengan toleransi Al...................................... 10
BAHAN DAN METODE
Tempat dan Waktu............................................................................. 13 Bahan ................................................................................................. 13 Metode Penelitian .............................................................................. 13
Perlakuan cekaman Al pada kultur cair ............................... 13 Isolasi RNA total ................................................................. 15 Sintesis cDNA Total ............................................................ 16 Ekpresi gen Gα dan gen PER .............................................. 16
HASIL DAN PEMBAHASAN
Pengaruh pH terhadap reduksi pertambahan panjang akar................ 18 Pengaruh Al terhadap reduksi pertambahan panjang akar................. 18 Isolasi RNA Total .............................................................................. 20 Sintesis cDNA Total .......................................................................... 20 Pengaruh Cekaman Aluminium Terhadap Ekspresi gen Heterotri- merik Gα ............................................................................................ 21 Pengaruh Cekaman pH dan Aluminium Terhadap Ekspresi Gen Peroksidase................................................................................. 23
SIMPULAN ..................................................................................................... 25 DAFTAR PUSTAKA ...................................................................................... 26 LAMPIRAN..................................................................................................... 32

DAFTAR TABEL No. Judul Halaman
1. Reduksi panjang akar oleh cekaman pH rendah ............................... 18 2. Reduksi perpanjangan akar tanaman kedelai kultivar Lumut yang mendapat perlakuan Al ..................................................................... 19 3. Ekspresi baku gen Gα pada tanaman kedelai kultivar Lumut dengan perlakuan pH..................................................................................... 22 4. Ekspresi baku gen Gα pada tanaman kedelai kultivar Lumut dengan perlakuan Al...................................................................................... 22 5. Ekspresi baku gen per pada tanaman kedelai kultivar Lumut dengan perlakuan Al...................................................................................... 23 6. Ekspresi baku gen per pada tanaman kedelai kultivar Lumut dengan perlakuan pH dan Al ......................................................................... 24
ii

DAFTAR GAMBAR No. Judul Halaman
1. RNA total akar kedelai cultivar lumut pada perlakuan pH 6, pH 4 dan pH 4 dengan cekaman 1.6 mM Al ................................................ 20 2. Hasil PCR Aktin yang berasal dari cetakan cDNA murni ................... 21 3. Hasil PCR Aktin yang cDNAnya terkontaminasi DNA genoma ........ 21 4. Ekspresi gen Gα dan aktin pada perlakuan pH 6, pH 4 dan pH 4+1.6 mM Al.................................................................................. 21 5. Ekspresi gen PER pada tanaman kedelai yang ditanam pada pH 6, pH 4 dan pH 4+1.6 mM Al ........................................................................... 23
iii

I. PENDAHULUAN
Latar Belakang
Kedelai (Glycine max (L.) Merr.) merupakan salah satu tanaman pangan yang
sangat penting di Indonesia. Biji kedelai dapat digunakan sebagai sumber protein
nabati dari bahan pangan dan pakan. Produksi kedelai dari tahun ke tahun cenderung
meningkat, namun belum dapat mengimbangi laju peningkatan permintaan dalam
negeri, sehingga import kedelai meningkat setiap tahun.
Salah satu faktor penyebab rendahnya produktivitas kedelai di Indonesia
adalah terbatasnya lahan tanam di pulau Jawa. Untuk mengatasi masalah ini adalah
dengan melakukan pembukaan lahan baru di luar pulau Jawa, sehingga akan terjadi
pergeseran penggunaan lahan tanam dari lahan subur ke lahan marginal. Sebagian
besar lahan marginal di Indonesia berupa lahan pasang surut, lahan salin, lahan
gambut dan lahan podzolik merah kuning. Lahan Podzolik Merah Kuning memiliki
pH rendah dan kandungan aluminium yang cukup tinggi dan miskin kandungan zat
hara tanah seperti Mg, Ca dan P. Jenis tanah tersebut luasnya mencapai 47,6 juta
hektar, dan banyak digunakan untuk program ekstensifikasi tanaman pangan termasuk
didalamnya tanaman kedelai (Notohadiprawiro 1983; Syarifuddin & Abdurahman
1993).
Aluminium diketahui sebagai salah satu faktor utama penyebab keracunan bagi
tanaman yang tumbuh di tanah yang bersifat masam, dengan potensi luasan di dunia
sekitar 1 X 109 hektar, mencakup daerah tropis dan sub tropis (Van Wambeke 1976;
Haug 1984; Moller et al. 1984). Al3+ merupakan bentuk paling toksik bagi tanaman
dengan gejala umumnya adalah pertumbuhan akar terhambat, akar menjadi pendek
dan menebal khususnya akar utama (Ryan et al. 1993, 1994; Sasaki et al. 1992, 1994;
Prihardi et al. 1995, Kochian 1995). Hal ini karena proses pembelahan dan
pemanjangan sel terganggu. Akibatnya, pertumbuhan dan perkembangan akar
terhambat, dan dalam jangka panjang akan mempengaruhi pertumbuhan dan
perkembangan bagian tajuk tanaman (Yamamoto et al. 1992).
Usaha peningkatan produksi kedelai pada lahan masam telah banyak
dilakukan, diantaranya dengan pengapuran (WidjajaAdhi 1985). Walaupun tidak
dipungkiri bahwa pengapuran mampu memperbaiki kondisi tanah masam, namun
dalam pelaksanaannya banyak ditemui hambatan seperti masalah distribusi kapur dan
1

aplikasinya di lapang. Maschner (1986) menyatakan bahwa toleransi yang tinggi
terhadap aluminium merupakan faktor kunci adaptasi tanaman pada tanah dengan pH
5.0. Tanaman-tanaman yang toleran terhadap tanah masam mampu meningkatkan pH
di daerah perakaran (Galvez & Clark 1991), mampu menyerap nitrat yang lebih besar
dan mampu membentuk komplek Al dengan asam organik (Christian-Waniger et al.
1992).
Penggunaan kedelai toleran pada lahan asam dengan kelarutan aluminium
tinggi merupakan salah satu alternatif dalam peningkatan produksi kedelai. Penelitian
untuk mendapatkan kultivar kedelai yang toleran terhadap tanah asam (Jusuf et al.
1999; Suharsono et al. 2003; 2006; 2007), serta isolasi gen-gen yang diduga diinduksi
oleh cekaman Al melalui penapisan differensial terhadap mRNA telah dilakukan
(Anwar et al. 2000). Penapisan differensial terhadap mRNA telah mendapatkan enam
klon cDNA tanaman kedelai kultivar Lumut yang peka terhadap Al dan satu klon
cDNA dari kultivar Slamet yang toleran Al (Anwar et al. 2000; Yuniati 2000).
Pembentukan pustaka genom kedelai dari kultivar Slamet (Suharsono 2002) dan
kultivar Lumut (Suharsono 2007) telah dilakukan. Penelitian Sopandie et al. (1996)
menunjukan bahwa genotipe kedelai yang toleran Al mengakumulasi lebih sedikit Al
dalam akar, dan mensintesis lebih banyak asam organik dibandingkan dengan genotipe
kedelai yang peka. Mekanisme signal transduksi di dalam sistem ketahanan tanaman
terhadap cekaman Al belum diketahui dengan jelas.
Protein G merupakan salah satu protein penting yang terlibat dalam
pensinyalan bagi sel dalam menanggapi perubahan lingkungan. Komponen jalur ini
terdiri dari reseptor terkait protein G yang berada pada permukaan membran dan
protein heterotrimerik G yang mentransduksi informasi dari reseptor protein G yang
teraktifasi ke efektor down stream seperti kanal ion, phospholipase C (Krauss 2001).
Protein G terlibat dalam jalur sinyal transduksi phosphoinositide pada mamalia dengan
melibatkan pemecahan lipid membran plasma phosphatidylinositol 4,5-biphosphate
(PtdInsP2) oleh phospholipase C (PLC) sehingga melepaskan inositol 1,4,5-
triphosphate (Ins[1,4,5]P3) ke sitoplasma dan DAG (Diacyl Glycerol) ke membran.
Selanjutnya Ins[1,4,5]P3 berasosiasi dengan reseptor membran (kanal Ca2+) untuk
mengeluarkan Ca2+ ke sitoplasma sehingga meningkatkan konsentrasi Ca2+ sitoplasma
dan DAG berasosiasi dengan protein kinase C (Krauss 2001; Cote & Crain 1993).
2

Al3+ dapat merusak metabolisme yang bergantung pada Ca dengan menjaga
konsentrasi Ca2+ sitoplasma diatas normal atau menghambat transien Ca2+ yang terjadi
secara bersama-sama (Delhaize & Ryan 1995). Menurut Yamamoto et al. (1997)
cekaman Al3+ dapat meningkatkan peroksidase phospholipid dan protein yang ada di
membran sel sehingga diduga Al dapat menyebabkan perubahan dalam proses
pemecahan phosphatidyl inositol 4,5-biphosphate (PIP2) menjadi DAG dan inositol
1,4,5-triphosphate (IP3). Jones & Kochian (1995) menyatakan bahwa cekaman Al
menghambat aktivitas PLC sehingga menghambat pemecahan PIP2 menjadi IP3 yang
akhirnya mempengaruhi influk ion Ca2+ ke dalam sel. Jalur sinyal transduksi berakhir
ke pengaturan satu atau lebih aktivitas gen atau pun pengatifan gen spesifik untuk
menanggapi sinyal dari luar.
Cekaman Al diketahui menginduksi beberapa gen yang juga berhubungan
dengan sistem pertahanan terhadap pathogen seperti peroksidase dan GST
(Richard et al 1998). Ekspresi gen-gen ini juga dipengaruhi oleh aktivitas species
oksigen aktif (ROS) H2O2 yang pertama terjadi pada tanaman pada saat tanaman
mengalami cekaman. H2O2 juga diketahui meningkat akibat aktivitas protein G subunit
alpha pada tanaman kedelai (Legendre et al. 1992).
Peroksidase termasuk ke dalam enzim golongan oksidoreduktase yaitu enzim
yang mengkatalis reaksi oksidasi-reduksi. Peroksidase tanaman tersebar dalam
jaringan tanaman yang ditemukan pada peroksisom. Enzim peroksidase berkaitan erat
dengan sejumlah proses fisiologi yang meliputi lignifikasi, penyembuhan luka,
oksidasi fenol, dan pertahanan terhadap patogen. Lignifikasi berperan sebagai
mekanisme pertahanan alami tanaman terhadap cekaman biotik dan abiotik. Lignin
merupakan biopolimer fenolik yang sintesisnya diinduksi oleh lingkungan dan
serangan patogen (Østergaard et al. 2000). Berdasarkan penelitian Cakmak & Horst
(1991) ditemukan bahwa aktivitas total peroksidase dan enzim anti-peroksidasi lainnya
pada ujung akar tanaman Glycine max meningkat selama perlakuan Al. Penelitian
Ezaki et al. (2000) menunjukkan bahwa peroksidase dan glutathione S-transferase
tidak bertindak langsung dalam menanggulangi cekaman Al dengan mereduksi
kandungan Al di dalam sel tanaman tetapi kemungkinan besar keduanya bertindak
memotong lipid peroksidasi di daerah membran sel sehingga membran sel ini dapat
menjaga masuknya ion Al ke dalam sitosol pada level rendah.
3

Oleh karena itu mekanisme sistem pertahanan tanaman terhadap cekaman Al
dengan melihat kemungkinan keterlibatan gen penyandi peroksidase (per) dan protein
G subunit α (Gα ) sangat penting untuk diketahui.
Tujuan Penelitian
Penelitian ini bertujuan untuk memperoleh informasi tentang ekspresi gen Gα
dan PER pada kedelai kultivar Lumut yang mendapat cekaman aluminium.
Hipotesis
Ekspresi Gα dan PER pada kedelai kultivar Lumut meningkat jika mendapat
cekaman Al dan pH 4
4

II. TINJAUAN PUSTAKA
Toksisitas dan Toleransi Aluminium pada Tanaman
Toksisitas Al. Aluminium bukanlah unsur esensial bagi pertumbuhan tanaman.
Saat kelarutan aluminium meningkat seiring dengan turunnya pH hingga di bawah 5
unsur ini menjadi penting untuk diperhatikan karena menjadi toksik bagi tanaman.
Bentuk-bentuk aluminium di dalam tanah dapat berupa ion trivalen yaitu Al(H2O)63+
atau disebut juga Al3+, bentuk hidroksida seperti Al(OH)+2, Al(OH)2+, Al(OH)3,
Al(OH)4-, atau berasosiasi dengan berbagai senyawa organik dan anorganik seperti
PO4-3, SO4
-2,F-, asam-asam organik, protein dan lipid (Delhaize & Ryan 1995). Al3+
merupakan bentuk yang paling toksik dan mendominasi di lahan asam di bawah pH
4.5 (Matsumoto 2000).
Tanah masam sendiri terjadi karena adanya pencucian kation-kation basa dari
tanah yang dipicu oleh praktek-praktek pertanian dan adanya hujan asam (Kennedy
1992). Oleh karena itu, untuk melihat pengaruh fitotoksik Al sebaiknya digunakan Al
dalam bentuk Al3+, pH media 4 dan kekuatan ioniknya rendah (Kinraide 1991)
Aluminium terlarut bereaksi dengan dinding dan membran sel akar serta
membatasi perluasan dinding sel sehingga menghentikan pemanjangan akar.
Terhentinya pemanjangan akar merupakan ciri utama dari toksisitas aluminium.
Jaringan akar merupakan bagian pertama dari tanaman yang mengalami keracunan
aluminium, terutama diujung akar sehingga mengalami pemendekan dan menebal.
Akar menjadi berwarna kecoklatan terutama pada akar utama serta terjadi
pertumbuhan akar lateral yang gemuk dan pendek dengan percabangan yang tidak
bagus (Sasaki et al. 1994; Ryan et al. 1993).
Selain itu Al3+ ini juga dapat memasuki sel melalui simplas setelah merusak
membran sel akar dan terkadang bereaksi dengan senyawa fosfor dan mengganggu
metabolisme fosfor pada tanaman. Akumulasi Al dalam sitoplasma memberi asumsi
bahwa toksisitas terjadi karena terbentuknya kompleks Al – ligan (Ryan et al. 1995).
Toksisitas aluminium melalui simplas disebabkan karena Al mengikat sangat kuat
terhadap senyawa donor O2 seperti Pi, nukleotida RNA, DNA, protein, asam
karboksilat, fosfolipid, asam poligalakturonat, heteropolisakarida, lipopolisakarida,
flavanoid, antosianin dan lain-lain (Haug 1984; Martin 1986). Konsentrasi Al yang
kecil saja dalam simplas berpotensi menjadi fitotoksik. Al3+ mengikat 107 kali lebih
5

kuat daripada Mg2+ terhadap ATP, sehingga jumlah Al3+ kurang dari nanomolar
sekalipun dapat menghambat Mg2+ pada situs P (Martin 1988). Aluminium juga
diduga menghambat proses pembelahan sel dan menghalangi metabolisme asam
nukleat (yaitu menghalangi reproduksi bahan genetik) pada tanaman (Helyar 1998).
Menurut Matsumoto (1991) Al yang berada dalam bentuk polimer (Al3+) memiliki
muatan positif yang besar serta memiliki banyak situs pengikatan. Polimer ini dapat
mengikat fosfat pada kedua utas DNA sehingga mengakibatkan gagalnya pemisahan
DNA utas ganda saat proses replikasi.
Toleransi Al. Pada prinsipnya ada dua mekanisme toleransi tanaman terhadap
cekaman Al menurut Taylor (1991), yaitu : pertama adalah mekanisme eksternal
yakni dengan mencegah Al masuk ke dalam simplas dan mencapai daerah metabolik
yang peka, sedangkan yang kedua adalah mekanisme internal yakni dengan
imobilisasi, kompartementasi atau detoksifikasi saat Al masuk ke dalam simplas
Mekanisme toleransi Al pada tanaman bervariasi baik antar maupun intra spesies.
Faktor genetik berperan penting dalam menentukan toleransi tersebut. Toleransi Al
pada gandum (Triticum aestivum) dikendalikan oleh sejumlah kecil gen dominan
mayor dan gen-gen ini telah dimanfaatkan di dalam program pemuliaan untuk merakit
kultivar yang toleran terhadap cekaman Al (Johnson et al. 1997).
Ada tiga jenis asam organik yang sering ditemukan dalam tanaman yang toleran
terhadap cekaman Al yaitu asam sitrat, asam oksalat dan asam malat. Sebagai contoh
respon tanaman terhadap cekaman Al adalah gandum yang mengeluarkan malat,
Snapbeans, jagung, Cassia toru dan kedelai yang melepas sitrat, Buckwheat
(Fagopyrum esculentum) yang mengeluarkan oksalat dan Triticale, Rapeseed, lobak,
oats dan Rye yang mengeluarkan malat dan sitrat. Mekanisme toleransi terhadap
cekaman Al dengan menggunakan asam organik ini dibagi ke dalam dua bentuk yaitu
detoksifikasi eksternal dan internal bahkan beberapa spesies tanaman menggunakan
kedua bentuk mekanisme tersebut.
Beberapa tanaman dapat mengakumulasi Al pada daun dan akarnya tanpa
menunjukkan gejala keracunan. Spesies tanaman toleran Al ini mempunyai
mekanisme untuk mengubah Al dalam bentuk non toksik di dalam tanaman yaitu
sebuah mekanisme yang membiarkan Al masuk ke dalam tanaman dan melewati
membran kemudian baru mengubahnya menjadi bentuk non toksik. Teh dan
6

hydrangea telah dikenal sebagai akumulator Al. Daun teh tua dapat mengakumulasi Al
hingga 30000 mg kg-1 berat kering (Matsumoto et al. 1976), dan akumulasi Al di daun
hydrangea mencapai di atas 3000 mg kg-1 (Ma et al. 1997). Melastoma malabathricum
dan Vaccinium macrocarpon yang beradaptasi baik pada pH rendah mengakumulasi
Al dalam level yang tinggi baik di daun maupun di akar (Osaki et al. 1997).
Meskipun banyak bukti yang menunjukkan keterkaitan asam organik dalam
mekanisme toleransi Al pada tanaman, ada beberapa spesies yang menunjukkan
mekanisme yang sama sekali tidak berkaitan dengan asam organik. Bachiaria
decumbans, salah satu jenis yang sangat toleran terhadap cekaman Al, tidak
mengeluarkan asam organik dalam merespon Al sehingga diyakini bahwa spesies ini
pasti memiliki mekanisme yang berbeda dalam menghadapi Al di level toksik pada
larutan tanah (Wenzl et al 2001). Arabidopsis mutan (alr1) meningkatkan pH yang
diinduksi oleh Al dengan segera di sekitar ujung akar yang dapat menurunkan aktivitas
Al3+ (Dengenhardt et al. 1998). Kultivar gandum yang sangat toleran Al (Atlas)
mempunyai mekanisme pelepasan fosfat sebagai mekanisme toleransi Al-nya (Pellet
et al. 1996). Berbeda dengan pelepasan malat, pelepasan posfat ini bersifat konstitutif
tanpa dipengaruhi adanya induksi Al untuk mengaktifkannya. Mekanisme lainnya
adalah sintesis protein spesifik yang diinduksi oleh cekaman oksidatif dan cekaman
Al. Cekaman Al menginduksi ekspresi beberapa gen seperti gen penyandi peroxidase,
glutathione S-transferase dan blue-copper protein. Overekspresi beberapa protein
tersebut pada Arabidopsis telah meningkatkan toleransi tanaman terhadap Al
sebagaimana peningkatan toleransi terhadap cekaman oksidatif (Ezaki et al. 2000).
Penemuan ini semakin menguatkan adanya mekanisme peningkatan toleransi Al
selain dari mekanisme pelepasan asam organik.
Menurut Richard et al. (1998) ekspresi dari gen GST, Peroxidase dan Blue
Copper binding protein dipengaruhi oleh aktifitas spesies oksigen aktif (AOS) H2O2
saat tanaman mengalami cekaman. Tanaman yang mampu melepaskan asam organik
sebagai mekanisme toleransi terhadap Al ternyata juga resisten terhadap cekaman
oksidatif.
Protein Heterotrimerik G sub unit α
Protein heterotrimerik G adalah protein peripheral membran plasma yang
menghadap ke permukaan interior sel (menghadap ke sitosol), merupakan reseptor
7

membran sel yang berfungsi sebagai mediator penyampai pesan/signal dari luar sel
(eksternal) ke molekul efektor sehingga menghasilkan respon intraseluler (Fujisawa et
al. 2001). Protein heterotrimerik G terdiri dari subunit α, β, γ (Fujisawa et al. 2001).
Masing-masing subunit tersebut terdiri dari 20 α, 6 β, 12 γ yang masing-masing berat
molekulnya adalah 42, 35, 10 kilo Dalton (kD) pada mamalia (Hamm 1998). Subunit α
memiliki 380 asam amino yang memiliki kesamaan pada tiap Gα (Bischoff et al.
1999).
Protein heterotrimerik G disebut protein G karena mengikat mononucleotide
GDP dan GTP. Subunit α merupakan subunit yang mengatur pertukaran GTP-GDP
pada mamalia (Fujisawa et al. 2001). Pada hewan/mamalia terdapat 4 famili subunit α
yaitu Gs (stimulatory) yang mengaktifkan enzim adenil siklase (cAMP), Gi (inhibitor)
yang menghambat adenil siklase, Gq mengaktifkan phospholipase Cβ (PLCβ), Gt
(bovin transducin) merangsang pembelahan cGMP (Ma 1994). Gα pada tanaman
memiliki homologi yang sama dengan mamalia. Ada kemungkinan βγ mempunyai
peranan secara langsung dalam meregulasi efektor dan interaksinya dengan reseptor
(Ma 1994).
Saat inaktif subunit α berikatan dengan GDP dan berasosiasi dengan βγ
membentuk kompleks. Ketika ligan terikat pada permukaan sel reseptor, reseptor
menjadi aktif dan mengkatalisis perubahan ikatan GDP pada subunit α menjadi GTP.
Hal tersebut menyebabkan terjadinya perubahan komformasi subunit α sehingga
akhirnya berpisah dengan βγ (disosiasi). Subunit α akan meregulasi efektor dengan
cara berikatan pada efektor dan mengaktifkan signal transduksi seperti pada adenilat
siklase. Protein heterotrimerik G kembali tidak aktif ketika GTP diubah menjadi GDP
dan subunit α kembali berasosiasi dengan βγ (Ma 1994).
Subunit α dari protein G terdapat pada plasma membran Arabidopsis (Weiss et
al. 1997), padi (Iwasaki et al. 1997) dan tembakau (Peskan & Muller 2000). Protein
heterotrimerik G sub unit α atau Gα mengaktifkan kanal Ca2+ pada membran plasma
tomat (Aharon et al. 1998), meningkatkan level IP3 kedelai (Legendre et al. 1993) dan
meningkatkan spesies oksigen aktif (AOS) H2O2 pada kultur sel kedelai (Legendre et
al. 1992). Protein heterotrimerik G berperan dalam meregulasi ketahanan terhadap
pathogen (Aharon et al. 1998; Beffa et al. 1995; Legendre et al. 1993), regulasi
8

lintasan biosintesis benzo phenathridine alkaloid (Mahady et al. 1998), dan regulasi
kanal K+ pada sel mesofil (Fairley-Grenot & Asmann 1991; Li & Asmann 1993).
Protein heterotrimerik G meregulasi banyak efektor yang ada dibawahnya
seperti adenilat siklase, PLC, dan efektor transducin (Ma 1994). PLC menghidrolisis
PIP2 menjadi 2 buah second messengers yaitu IP3 dan DAG. IP3 dapat mengikat
reseptor membran seperti kanal Ca2+, melepas Ca2+ dari reticulum endoplasmik ke
dalam sitosol sehingga level Ca2+ meningkat. Peningkatan Ca2+ di sitoplasmik
memproduksi protein kinase C (PKC), DAG berperan dalam mengaktifkan protein
kinase C (Mc Laughlin et al. 1996; Krauss 2001 ).
Membran plasma sel akar yang kaya akan fosfatase yang berbentuk pospolipid
merupakan target utama Al3+ (Matsumoto 2000). Al3+ berinteraksi dengan lipid
membran plasma dan dengan enzim yang mengikat metal (Jones & Kochian 1996).
Al3+ dapat menyebabkan sitotoksik yang kronik pada tanaman, hewan dan
mikroorganisme, sehingga menyebabkan penghambatan perpanjangan akar
(elongation). Apex akar (tudung akar, sel meristem dan zone perpanjangan)
mengakumulasi lebih banyak Al, serta mengalami kerusakan yang lebih parah
dibandingkan dengan jaringan akar yang telah dewasa dan berperan dalam mekanisme
respon terhadap Al (Matsumoto 2000; Delhaize & Ryan 1995). Mekanisme toleran
terhadap Al pada gandum terjadi dengan cara mengkelat Al3+, imobilisasi Al3+ pada
dinding sel, meningkatkan pH disekitar akar dan mengeluarkan Al dari sitoplasma
(Taylor 1991). Al menghambat pertumbuhan dan perpanjangan akar squash
(Curcubita maxima putch) dengan mengubah metabolisme polisakarida dinding sel di
daerah non elongation sama seperti di daerah elongation (Van et al. 1994).
Genotipe kedelai toleran mengakumulasi Al dalam jumlah rendah pada sel
meristem dan sel diferensiasi pada ujung akar dan dinding sel (Silva et al. 2000).
Akibat cekaman Al tanaman mengeluarkan asam organik dari apex akar ke rhizosfer
yang berfungsi untuk mengkelat ligan Al, seperti eksudasi sitrat (Pineros et al. 2002),
malat pada gandum (Triticum aestivum) (Ryan et al. 1995; Tang et al. 2002), sitrat
pada jagung (Zea mays) (Pellet et al. 1995), snapbean (Miyasaka et al. 1991), asam
malat pada gandum (Delhaize & Ryan 1995), oksalat pada buckwheat (Ma et al.
1997).
9

Cekaman aluminium juga diketahui menginduksi beberapa gen yang
berhubungan dengan sistem pertahanan terhadap pathogen (defense-response) seperti
peroksidase, GST dan blue copper binding protein (AtBCB) pada Arabidopsis
thaliana (Richard et al. 1998).
Gen yang berhubungan dengan toleransi Al
Pada kedelai, beberapa gen yang ekspresinya diinduksi oleh Al telah diisolasi.
Pada kultivar yang peka terhadap cekaman Al yaitu Lumut, enam klon cDNA yang
ekspresinya diinduksi Al telah diisolasi (Anwar et al. 2000). Dari kultivar toleran yaitu
Slamet, menggunakan teknik penapisan diferensial terhadap mRNA dari akar tanaman
yang mendapat cekaman Al telah diisolasi satu klon (Yuniati 2000).
Untuk menganalisis gen-gen tersebut lebih lanjut, pustaka genom kedelai
kultivar Lumut dan Slamet telah dikonstruksi (Suharsono 2002, 2007) dalam fage λ
sebagai vektor.
Salah satu gen yang penting dalam sistem toleransi tumbuhan terhadap
cekaman Al adalah peroksidase. Peroksidase termasuk ke dalam enzim golongan
oksidoreduktase yaitu enzim yang mengkatalis reaksi oksidasi-reduksi. Molekul-
molekul toksik seperti superoksida dan radikal hidroksida terkumpul di dalam sel
bersamaan dengan adanya oksigen. Molekul-molekul toksik ini merupakan produk
intermediate dari respirasi aerobik namun dibatasi jumlahnya di dalam sel oleh
sejumlah enzim. Superoksida dipecahkan oleh SOD dimana dalam proses tersebut
dihasilkan hidrogen peroksida (H2O2) yang kemudian diuraikan oleh peroksidase.
Peroksidase memecahkan H2O2 menjadi air ketika mengoksidasi sejumlah substrat.
Dengan demikian peroksidase adalah enzim golongan oksidoreduktase yang
menggunakan H2O2 sebagai akseptor elektron untuk mengkatalis berbagai reaksi
oksidatif. Peroksidase juga membutuhkan kofaktor untuk melakukan aktivitas
enzimatisnya yaitu heme sehingga peroksidase disebut juga heme dependent
peroxidase. Heme adalah kompleks antara ion besi dan molekul protoporphyrin IX
(Ferriprotoporphyrin IX).
Berdasarkan kemiripan sekuen dan strukturnya, heme dependent peroxidase
dibedakan ke dalam 2 famili yaitu : 1) peroksidase hewan dan 2) peroksidase tanaman,
fungi dan bakteri. Famili peroksidase tanaman terdiri dari 3 kelas yang dibedakan
berdasar struktur dan fungsinya. Kelas I terdiri dari peroksidase intraseluler yang
10

terdapat pada fungi, tanaman, archaea dan bakteri, disebut peroksidase-katalase yang
menunjukkan aktivitas katalase dan peroksidase dalam spektrum yang luas dan
tergantung pada konsentrasi hidrogen peroksida yang mantap. Kelas II terdiri dari
ligninase dan peroksidase ekstraseluler fungi lainnya, sedangkan kelas III terdiri dari
peroksidase ekstraseluler klasik tanaman seperti horseradish peroksidase.
Peroksidase tanaman tersebar dalam jaringan tanaman yang ditemukan pada
peroksisom. Enzim peroksidase berkaitan erat dengan sejumlah proses fisiologi yang
meliputi lignifikasi, penyembuhan luka, oksidasi fenol, dan pertahanan terhadap
patogen.
Lignifikasi berperan sebagai mekanisme pertahanan alami tanaman terhadap
cekaman biotik dan abiotik. Lignin merupakan biopolimer fenolik yang sintesisnya
diinduksi oleh lingkungan dan serangan patogen (Østergaard et al. 2000). Monomer
lignin berasal dari tiga hydroxycinnamyl alkohol atau monolignols yaitu p-coumaryl,
coniferil dan sinaphyl. Ketiga monolignol ini menghasilkan monolignin p-
hidroksifenil (H), guasil (G) dan syringil (S) yang bergabung membentuk polimer
lignin. Polimerisasi dehidrogenase dilakukan dengan bantuan peroksidase tanaman,
laccases (oksidoreduktase) atau keduanya. Peroksidase dapat mengoksidasi spektrum
fenolik yang luas termasuk proses lignifikasi monomer secara in vitro (Dunford,
1991).
Berdasarkan penelitian yang dilakukan Hiraga et al. (2001) tingginya tingkat
aktivitas peroksidase pada tanaman berkaitan erat dengan peningkatan resistensi
terhadap cekaman abiotik, termasuk cekaman oksidatif. Cekaman oksidatif disebabkan
oleh berbagai faktor lingkungan diantaranya cekaman UV, serangan pathogen
(hypersensitive reaction), herbisida dan kekurangan oksigen (Blokhina et al. 2002).
Aktivitas kelompok oksigen aktif (ROS) merupakan salah satu mekanisme yang
muncul pertama kali pada tanaman untuk mengatasi cekaman oksidatif khususnya
untuk reoksigenasi. Pada ROS ini hidrogen peroksida (H2O2) dan superoksida (O2-) di
hasilkan di sejumlah reaksi selular yang dikatalis oleh beberapa enzim seperti
lipoksigenase, peroksidase, NADPH oksidase dan xanthine oksidase. Peroksidase dan
katalase berperan penting dalam regulasi konsentrasi ROS di dalam sel dengan
aktivasi dan deaktivasi H2O2
11

Peroksidase anionik tembakau (NtPox) dan glutathione S-transferase (parB)
merupakan protein intraseluler yang digunakan untuk mendetoksifikasi kelompok
oksigen reaktif (Reactive Oxygen Species). Berdasarkan penelitian Cakmak & Horst
(1991) ditemukan bahwa aktivitas total peroksidase dan enzim anti-peroksidasi lainnya
pada ujung akar tanaman Glycine max meningkat selama perlakuan Al. Penelitian
Ezaki et al. (2000) menunjukkan bahwa peroksidase dan glutathione S-transferase
tidak bertindak langsung dalam menanggulangi cekaman Al dengan mereduksi
kandungan Al di dalam sel tanaman tetapi kemungkinan besar keduanya bertindak
memotong lipid peroksidasi di daerah membran sel sehingga membran sel ini dapat
menjaga masuknya ion Al ke dalam sitosol pada level rendah.
12

III. BAHAN DAN METODE
Tempat dan Waktu
Penelitian dilakukan di Laboratorium BIORIN (Biotechnology Research
Indonesian - The Netherlands) Pusat Penelitian Sumberdaya Hayati dan Bioteknologi
IPB. Penelitian dimulai dari bulan Januari 2005 sampai dengan bulan Januari 2007.
Bahan
Bahan tanaman yang digunakan adalah kedelai kultivar Lumut yang peka
terhadap cekaman aluminium (Anwar et al. 1999). Cekaman Al menggunakan AlCl3.
Primer forward (F) (5’CCCAAGCTTGGTACCCGCGTCTGTTGACTGGCAGG3’)
dan Primer reverse (R)
(5’CCCTCTAGACTCGAGTGGAGATGGTGCTGTTGGTCC3’) digunakan untuk
amplifikasi cDNA dari gen penyandi peroksidase. Primer (F) terletak pada 111
nukleotida sebelum kodon awal (5’GCTTCACACTTCACACTTAACACT3’) dan
primer (R) terletak pada 114 nukleotida sesudah kodon akhir
(5’ATATTGTTGTATACCTGACCTC3’) yang didesain dari kedelai, yaitu SGAI
(nomor aksesi L27418), digunakan untuk amplifikasi cDNA dari gen penyandi Gα.
Primer F tepat pada kodon awal dari ekson 1 (5’ATGGCAGATGCCGAGGATAT3’)
dan primer R tepat pada daerah ekson 2 (5’CAGTTGTGCGACCACTTGCA3’),
didesain dari kedelai (nomor aksesi V00450), digunakan untuk amplifikasi β-aktin
Metode Penelitian
Analisis ekspresi gen Gα dan PER dilakukan melalui beberapa tahapan, yaitu:
(1) Perlakuan cekaman pada kultur cair, (2) Isolasi RNA total, (3) Sintesis cDNA total,
dan (4) Analisis ekspresi gen Gα dan PER.
(1) Perlakuan cekaman Al pada kultur cair
Langkah awal adalah menyeleksi biji kedelai kultivar Lumut yang memiliki
ukuran yang sama kemudian benih dikecambahkan selama dua hari. Benih
dikecambahkan dalam kertas merang, disimpan dalam ruang gelap dengan
kelembaban yang tinggi. Setelah berkecambah, kecambah ditanam di atas wadah
13

ukuran 20 cm x 30 cm yang berlubang. Wadah tersebut diletakkan di atas bak plastik
ukuran 25 cm x 35 cm x 15 cm yang telah berisi media cair pH 6 sebagai media
tanamnya, dan diberi aerasi dengan menempatkan aerator sebanyak 4 buah tiap wadah
untuk menjaga ketersediaan oksigen. Kecambah diletakkan sedemikian rupa sehingga
ujung akar menyentuh media cair. Komposisi media tanam adalah 0.375 mM
Ca(NO3)2. 4H2O, 0.2 µM CuSO4.5H2O, 0.25 mM NH4NO3, 1 µM ZnSO4.7H2O, 0.1
mM Mg SO4 . 7H2O, 5 µM H3BO3, 0.1mM KH2PO4, 1 µM (NH4)6Mo7O24.4H2O, 5
µM MnSO4.H2O, 5 µM Fe-EDTA, dengan pH 6 (Anwar 2000). Kecambah
ditumbuhkan pada media ini selama 2 hari
Tahapan pada hari berikutnya adalah pemberian perlakuan. Perlakuan yang
diberikan ada dua macam yaitu perlakuan pH dan perlakuan Al. Bak pertama
digunakan untuk pH 6 yang merupakan kontrol perlakuan pH, bak kedua untuk pH 4.
Bak kedua yang berisi media dengan pH 4 juga digunakan sebagai kontrol perlakuan
Al. Bak ketiga untuk perlakuan pH 4 dengan cekaman 1,2 mM Al dan bak keempat
untuk perlakuan pH 4 dengan cekaman 1,6 mM Al. Perlakuan dilakukan selama 3x24
jam, dengan media diganti setiap 24 jam. Pengamatan panjang akar utama dilakukan
terhadap 10 sampel yang diambil secara acak, dengan mengukur akar dari pangkal
batang sampai dengan ujung akar. Percobaan dilakukan dengan dua ulangan. Pada saat
pengamatan, sekitar 0,3 cm ujung akar utama diambil, dibungkus dengan aluminium
foil lalu difiksasi di dalam nitrogen cair dan disimpan di freezer suhu -40˚C. Bahan
tanaman ini nantinya digunakan untuk isolasi RNA total.
Pemilihan konsentrasi Al yang optimum dan lama cekaman ditentukan
berdasarkan persentase selisih perpanjangan akar pada setiap jam perlakuan.
Konsentrasi Al dan pH serta lama cekaman yang optimum menghambat pertumbuhan
akar digunakan untuk uji ekspresi gen Gα dan gen PER. Reduksi atau stimulasi
perpanjangan akar dihitung berdasarkan nilai perbandingan pertambahan panjang akar
dari perlakuan terhadap pertambahan panjang akar dari kontrol. Kontrol pada
percobaan ini adalah tanaman yang ditumbuhkan pada pH 4,0 untuk perlakuan Al dan
pH 6 sebagai kontrol perlakuan pH. Reduksi perpanjangan akar dihitung dengan
rumus:
100)()()( ×= −
−−−YtoYti
XtoXtiYtoYtiRPA
14

atau
100×= −PPAy
PPAxPPAyRPA
RPA : Reduksi panjang akar
Yti : Panjang akar dari tanaman kontrol pada waktu ti
Yto : Panjang akar dari tanaman kontrol pada waktu to
Xti : Panjang akar dari tanaman yang diperlakukan pada waktu ti
Xto : Panjang akar dari tanaman yang diperlakukan pada waktu to
PPAy : Pertambahan panjang akar tanaman kontrol
PPAx : Pertambahan panjang akar tanaman perlakuan
Nilai RPA positif menunjukkan bahwa perlakuan menyebabkan reduksi pertambahan
panjang akar bila dibandingkan dengan kontrol, yaitu tanaman yang ditumbuhkan pada
pH 4,0. Nilai RPA negatif menunjukkan bahwa perlakuan menyebabkan stimulasi
pertambahan panjang akar dibandingkan dengan kontrol.
(2) Isolasi RNA total
Sekitar 50-100 mg ujung akar kedelai yang telah tersimpan dalam aluminum
foil pada suhu -400C, diberi nitrogen cair langsung digerus dengan menggunakan
mortar sampai halus berbentuk bubuk. Bubuk dicampurkan 800 µl TRIzol. Suspensi
sel dipindahkan ke dalam ependorf dan diinkubasikan pada suhu ruang selama kurang
lebih 5 menit. Ke dalam ependorf, 200 µl kloroform dimasukkan dan suspensi sel
divortex sampai tercampur. Campuran diinkubasikan pada suhu ruang selama 3 menit.
Selanjutnya ependorf tersebut disentrifugasi dengan kecepatan 9000 rpm (Jouan BR4i)
dengan suhu 6˚C selama 15 menit. Cairan bagian atas diambil sebanyak minimal 60%
dari volume TRIzol. Supernatan tersebut dipindahkan ke dalam ependorf baru, dan
ditambahkan isopropil alkohol lalu diinkubasikan dalam suhu ruang selama 10 menit.
Setelah itu ependorf tersebut disentrifugasi dengan kecepatan 9000 rpm selama 10
menit dengan suhu 6˚C. Supernatan dari hasil sentrifugasi dibuang, dan endapannya
diambil. Kemudian ditambah dengan etanol 75% lalu divortex. Ependorf kembali
disentrifugasi dengan kecepatan 5700 rpm selama 5 menit dengan suhu 6˚C. Etanol
75% dibuang, endapan dikeringkan dengan menggunakan vakum. Setelah kering
endapan disuspensikan dalam 30 µl DEPC 0.1% (v/v).
15

(3) Sintesis cDNA Total
Sintesis cDNA dilakukan dengan transkripsi balik menggunakan RNA total
sebagai cetakan dengan metode Suharsono et al. (2002). Sebanyak 500 ng RNA total
dicampur dengan 4 µl buffer (5x), 2 µl 2 mM dNTPmix, 2 µl 0.1 M dTT, 10 pmol
primer oligo dT, 1 unit enzim reverse transcriptase (RT) dan DEPC hingga volume
akhir reaksi 20 µl. Kondisi reaksi transkripsi balik adalah 10 menit suhu 30˚C, 50
menit suhu 42˚C, 5 menit suhu 95˚C˚.
Evaluasi keberhasilan sintesis cDNA total dilakukan melalui PCR dengan
menggunakan primer β-aktin. PCR β-aktin dilakukan dengan mencampur 2 µl cDNA
total, 2 µl buffer (10x), 1 µl 2 mM dNTPmix, 0,8 µl 25 mM MgCl2, 1unit enzim taq
DNA polimerase, 10 pmol primer aktin forward (F), 10 pmol primer aktin reverse
(R), 0,4 µl DMSO digenapkan dengan ddH2O hingga 20 µl. Kondisi yang digunakan
adalah Pra-PCR 95˚C 5 menit, denaturasi 94˚C 30 detik, annealing 55˚C 30 detik,
ekstensi 72˚C 1 menit 40 detik, siklus diulangi sebanyak 30 kali, pasca-PCR 72˚C 5
menit. Apabila cDNA yang disintesis adalah murni yang tidak terkontaminasi DNA
genom, PCR menghasilkan amplifikasi berukuran 450 pb. Apabila terkontaminsi DNA
genom, maka hasil PCR berukuran 450 pb dan 550 pb karena cetakan cDNA
menghasilkan 450 pb dan cetakan DNA genom menghasilkan 550 pb yang meliputi
daerah ekson 1, intron dan ekson 2. Selain untuk melihat keberhasilan sintesis cDNA
total dari kontaminasi DNA genom, PCR β-aktin juga digunakan untuk menyetarakan
konsentrasi cDNA pada berbagai perlakuan. Untuk mengetahui ukuran PCR aktin,
dilakukan elektroforesis pada gel agarosa TAE 1x (0,04 M Tris-acetat, 0,001 M
EDTA).
(4) Ekspresi gen Gα dan gen PER
Analisis ekspresi gen dilakukan pada tanaman kedelai yang mendapat
perlakuan pH dan perlakuan Al secara terpisah. Perlakuan pH yang diberikan adalah
pH 4 dan pH 6 sebagai kontrol. Perlakuan Al menggunakan pH 4 tanpa penambahan
Al sebagai kontrol. Percobaan dilakukan dengan dua ulangan.
Analisis ekspresi gen Gα dan gen PER dilakukan dengan cara mengamplifikasi
gen spesifik tersebut dengan menggunakan cDNA total sebagai cetakannya. Langkah
dalam mencampur bahan untuk PCR gen Gα dan gen PER sama dengan PCR aktin,
16

yang membedakan hanya primer yang disesuaikan dengan gen yang dianalisis.
Kondisi PCR gen Gα sama dengan kondisi PCR aktin, sedangkan untuk kondisi PCR
gen PER yaitu Pra-PCR 94˚C 2 menit, denaturasi 92˚C selama 30 detik,
annealing/penempelan 55˚C selama 30 detik, ekstensi 75˚C selama 1 menit, siklus
diulang sebanyak 30 kali, dan pasca PCR 72˚C selama 5 menit. Analisis ekspresi
dilakukan dengan membandingkan intensitas cahaya pita hasil PCR gen Gα dan PER
terhadap kontrol aktin dengan menggunakan perangkat lunak Digi Doc-it.
Agar dapat diperbandingkan ekspresi gen sasaran tertentu pada waktu yang
sama pada berbagai perlakuan, maka ekspresi gen sasaran tertentu harus dibakukan.
Pembakuan ekspresi gen sasaran tertentu dilakukan dengan membandingkan ekspresi
gen sasaran dengan gen aktin pada waktu dan perlakuan yang sama. Oleh sebab itu
ekspresi gen sasaran Gα, dan gen PER dibakukan dengan menggunakan rumus :
IAptIXptEBXpt =
EBXpt : Ekspresi baku gen x pada perlakuan p waktu t
IXpt : Intensitas hasil PCR gen x pada perlakuan p waktu t
IApt : Intensitas hasil PCR gen aktin pada perlakuan p waktu t
x : Gα dan PER
t : 0, 8, 24, 48, atau 72 jam perlakuan cekaman
17
IV. HASIL DAN PEMBAHASAN
Pengaruh pH terhadap reduksi pertambahan panjang akar
Fenotip reduksi pertambahan panjang akar pada perlakuan pH 4 dibandingkan
dengan kontrol (pH6) secara visual tidak terlalu terlihat perbedaan namun pada
perlakuan pH 4 + Al baik 1,2 mM maupun 1,6 mM terlihat sangat jelas (Lampiran 3).
Pengaruh pH 4 terhadap reduksi panjang akar menggunakan kontrol pH 6.
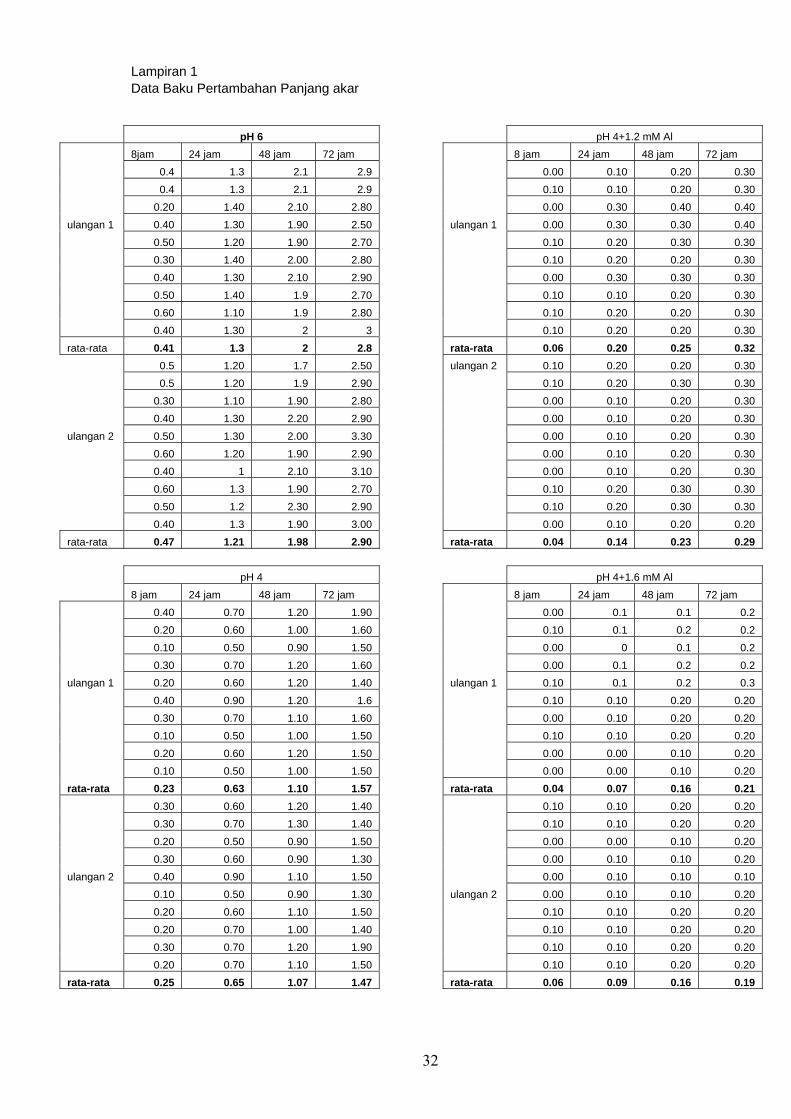
Hasil analisis reduksi panjang akar disajikan pada Tabel 1.
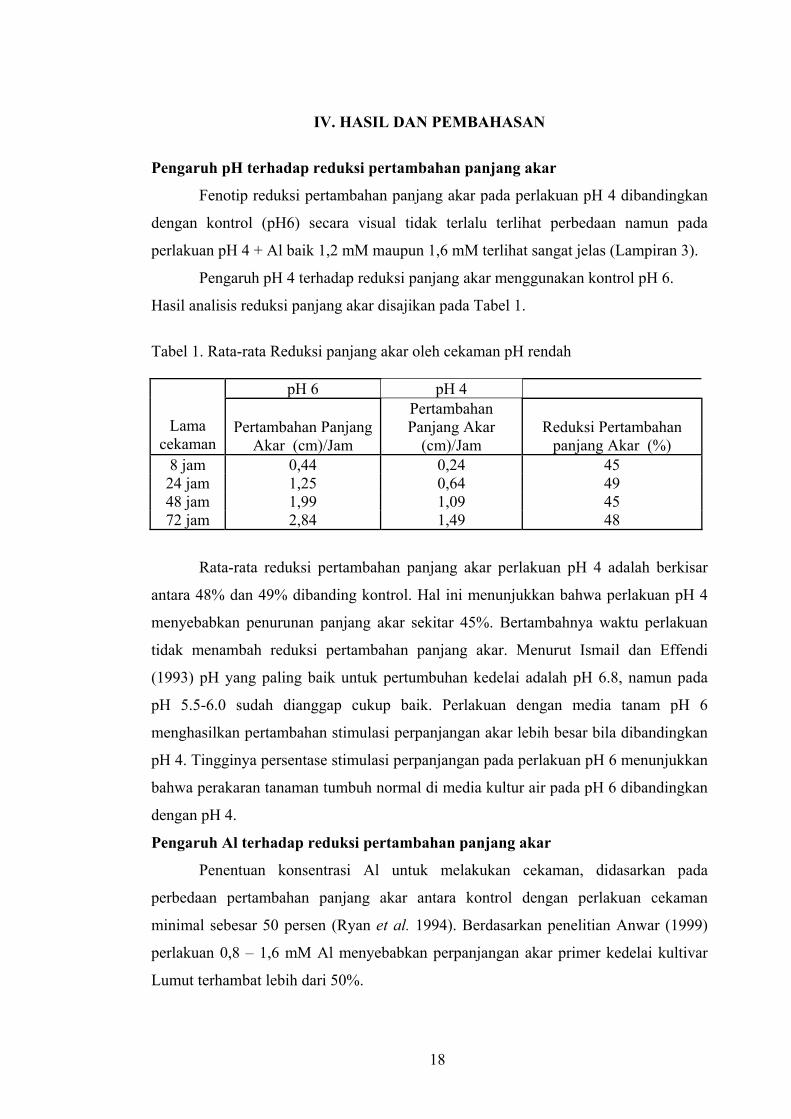
Tabel 1. Rata-rata Reduksi panjang akar oleh cekaman pH rendah
pH 6 pH 4
Lama cekaman
Pertambahan Panjang Akar (cm)/Jam
Pertambahan Panjang Akar
(cm)/Jam Reduksi Pertambahan
panjang Akar (%) 8 jam 0,44 0,24 45 24 jam 1,25 0,64 49 48 jam 1,99 1,09 45 72 jam 2,84 1,49 48
Rata-rata reduksi pertambahan panjang akar perlakuan pH 4 adalah berkisar
antara 48% dan 49% dibanding kontrol. Hal ini menunjukkan bahwa perlakuan pH 4
menyebabkan penurunan panjang akar sekitar 45%. Bertambahnya waktu perlakuan
tidak menambah reduksi pertambahan panjang akar. Menurut Ismail dan Effendi
(1993) pH yang paling baik untuk pertumbuhan kedelai adalah pH 6.8, namun pada
pH 5.5-6.0 sudah dianggap cukup baik. Perlakuan dengan media tanam pH 6
menghasilkan pertambahan stimulasi perpanjangan akar lebih besar bila dibandingkan
pH 4. Tingginya persentase stimulasi perpanjangan pada perlakuan pH 6 menunjukkan
bahwa perakaran tanaman tumbuh normal di media kultur air pada pH 6 dibandingkan
dengan pH 4.
Pengaruh Al terhadap reduksi pertambahan panjang akar
Penentuan konsentrasi Al untuk melakukan cekaman, didasarkan pada
perbedaan pertambahan panjang akar antara kontrol dengan perlakuan cekaman
minimal sebesar 50 persen (Ryan et al. 1994). Berdasarkan penelitian Anwar (1999)
perlakuan 0,8 – 1,6 mM Al menyebabkan perpanjangan akar primer kedelai kultivar
Lumut terhambat lebih dari 50%.
18
Oleh sebab itu, konsentrasi 1,2 mM Al dan 1,6 mM Al digunakan untuk
melakukan cekaman dan pH 4 sebagai kontrol. Hasil reduksi pertambahan panjang
akar dari tanaman kedelai kultivar Lumut yang mendapat cekaman Al disajikan pada
Tabel 2.
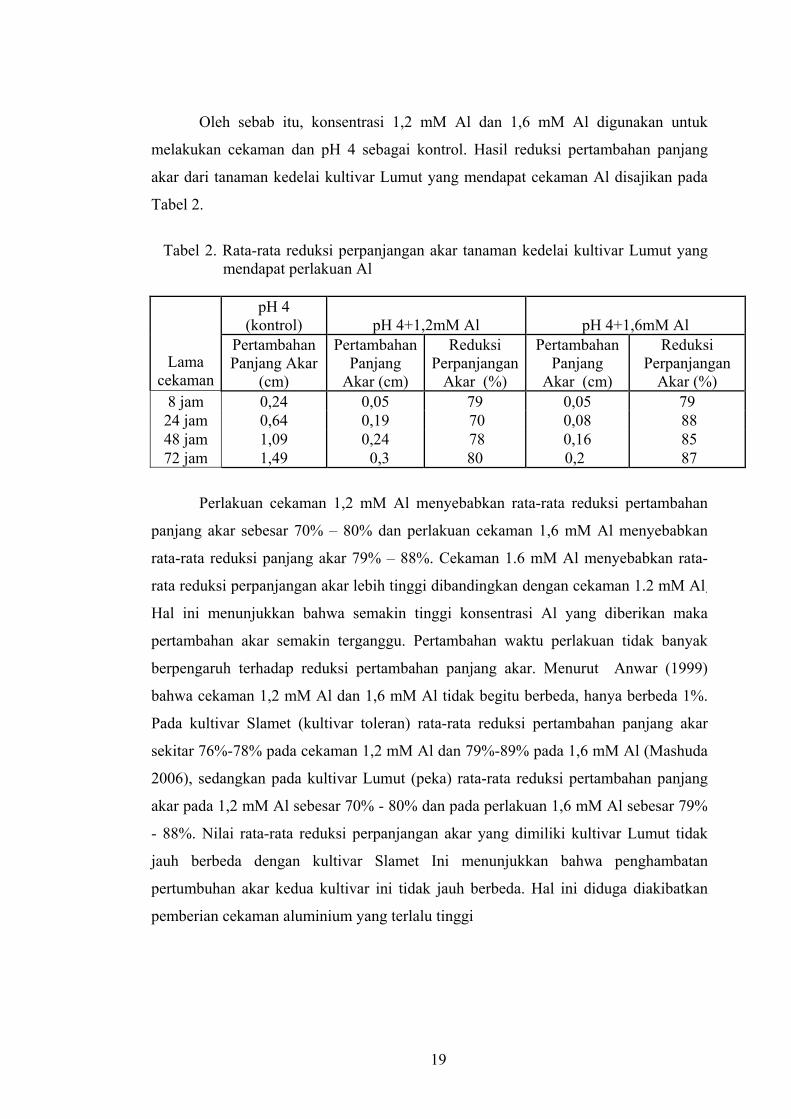
Tabel 2. Rata-rata reduksi perpanjangan akar tanaman kedelai kultivar Lumut yang
mendapat perlakuan Al
pH 4 (kontrol) pH 4+1,2mM Al pH 4+1,6mM Al
Lama cekaman
Pertambahan Panjang Akar
(cm)
Pertambahan Panjang
Akar (cm)
Reduksi Perpanjangan
Akar (%)
Pertambahan Panjang
Akar (cm)
Reduksi Perpanjangan
Akar (%) 8 jam 0,24 0,05 79 0,05 79 24 jam 0,64 0,19 70 0,08 88 48 jam 1,09 0,24 78 0,16 85 72 jam 1,49 0,3 80 0,2 87
Perlakuan cekaman 1,2 mM Al menyebabkan rata-rata reduksi pertambahan
panjang akar sebesar 70% – 80% dan perlakuan cekaman 1,6 mM Al menyebabkan
rata-rata reduksi panjang akar 79% – 88%. Cekaman 1.6 mM Al menyebabkan rata-
rata reduksi perpanjangan akar lebih tinggi dibandingkan dengan cekaman 1.2 mM Al.
Hal ini menunjukkan bahwa semakin tinggi konsentrasi Al yang diberikan maka
pertambahan akar semakin terganggu. Pertambahan waktu perlakuan tidak banyak
berpengaruh terhadap reduksi pertambahan panjang akar. Menurut Anwar (1999)
bahwa cekaman 1,2 mM Al dan 1,6 mM Al tidak begitu berbeda, hanya berbeda 1%.
Pada kultivar Slamet (kultivar toleran) rata-rata reduksi pertambahan panjang akar
sekitar 76%-78% pada cekaman 1,2 mM Al dan 79%-89% pada 1,6 mM Al (Mashuda
2006), sedangkan pada kultivar Lumut (peka) rata-rata reduksi pertambahan panjang
akar pada 1,2 mM Al sebesar 70% - 80% dan pada perlakuan 1,6 mM Al sebesar 79%
- 88%. Nilai rata-rata reduksi perpanjangan akar yang dimiliki kultivar Lumut tidak
jauh berbeda dengan kultivar Slamet Ini menunjukkan bahwa penghambatan
pertumbuhan akar kedua kultivar ini tidak jauh berbeda. Hal ini diduga diakibatkan
pemberian cekaman aluminium yang terlalu tinggi
19

Agar pengaruh cekaman memberikan hasil yang nyata, maka respon tanaman
yang mengalami cekaman harus menampakkan perbedaan yang cukup jelas.
Berdasarkan hasil yang didapat, dari cekaman 1.6 mM Al menghasilkan rata-rata
reduksi perpanjangan akar (RPA) yang lebih tinggi dibandingkan pada cekaman 1.2
mM Al, sehingga analisis ekspresi gen pada tahap berikutnya hanya menggunakan
cekaman Al pada konsentrasi 1.6 mM Al.
Isolasi RNA Total
Isolasi RNA total telah berhasil dilakukan. RNA diisolasi dari ujung akar
tanaman kedelai kultivar Lumut yang mendapat perlakuan cekaman pH dan Al.
Integritas RNA total yang diisolasi pada penelitian ini adalah baik. Hal ini ditandai
oleh dua pita rRNA yang dominan (28S dan 18S) yang utuh (Gambar 1). Karena
mRNA terdapat bersama-sama dengan rRNA didalam RNA total, maka bilamana
rRNA tersebut utuh maka mRNA juga utuh. RNA total ini kemudian digunakan untuk
cetakan dalam sintesis cDNA total. pH 6 pH 4 pH 4 + 1,6 mM Al 0 J 8 J 24 J 48 J 72 J 8 J 24 J 48 J 72J 8J 24J 48J 72J
28 s 18 s
Gambar 1. RNA total akar kedelai kultivar Lumut pada perlakuan pH 6, pH 4 dan
pH 4 dengan cekaman 1,6mM Al.
Síntesis cDNA Total
cDNA total telah berhasil diisolasi dengan menggunakan RNA total sebagai
cetakannya. Kemurnian cDNA dianalisis dengan PCR dengan menggunakan primer
untuk ekson 1 – ekson 2 dari β-aktin. Aktin juga digunakan sebagai kontrol internal
karena setiap sel mempunyai aktin dan ekspresinya bersifat konstitutif. PCR dengan
primer untuk ekson 1 – ekson 2 dari β-aktin menghasilkan satu pita DNA berukuran
450 pb (Gambar 2) yang menunjukkan bahwa cDNA total adalah murni yang tidak
terkontaminasi oleh DNA genom. Adanya kontaminasi DNA genom menyebabkan
hasil amplifikasi β-aktin menghasilkan 2 pita yaitu yang berukuran sekitar 450 pb dan
550 pb (Gambar 3) karena DNA genom mengandung intron diantara ekson 1 dan
20

ekson 2 yang berukuran sekitar 100 pb. cDNA total yang murni digunakan sebagai
cetakan untuk analisis ekspresi gen melalui PCR.
pH 6 pH 4 pH 4 + 1,6 mM Al 0 J 8 J 24 J 48 J 72 J 8 J 24 J 48 J 72J 8J 24J 48J 72J
450 pb
Gambar 2. Hasil PCR Aktin yang berasal dari cetakan cDNA murni.
1 2
450 pb
550 pb
Gambar 3. Hasil PCR β-aktin yang cDNAnya terkontaminasi DNA genom (1), dan
cDNA murni (2).
Pengaruh Cekaman Aluminium Terhadap Ekspresi Gen Heterotrimerik Gα.
Untuk membedakan pengaruh pH dan pengaruh Al terhadap ekspresi gen,
maka kontrol yang digunakan untuk analisis gen pada perlakuan pH berbeda dengan
perlakuan Al. Perlakuan pH menggunakan kontrol pH 6 sedangkan perlakuan Al
menggunakan kontrol pH 4 karena perlakuan Al tidak dapat dilakukan pada pH 6.
Pita hasil PCR untuk melihat hasil ekspresi gen Gα disajikan pada Gambar 4
dan pengukuran intensitas pita disajikan pada Tabel 3 dan Tabel 4. Dari hasil
pengukuran intensitas pita menunjukkan adanya perbedaan tingkat ekspresi gen. Hal
ini menunjukkan bahwa adanya perbedaan respon gen terhadap berbagai cekaman
yang dicobakan. pH 6 pH 4 pH 4 + 1,6 mM Al 0 J 8 J 24 J 48 J 72 J 8 J 24 J 48 J 72J 8J 24J 48J 72J
β-aktin 450 pb
Gα 1380 pb
Gambar 4. Ekspresi gen Gα dan aktin pada perlakuan pH 6, pH 4, dan pH 4+1,6mM Al.
21
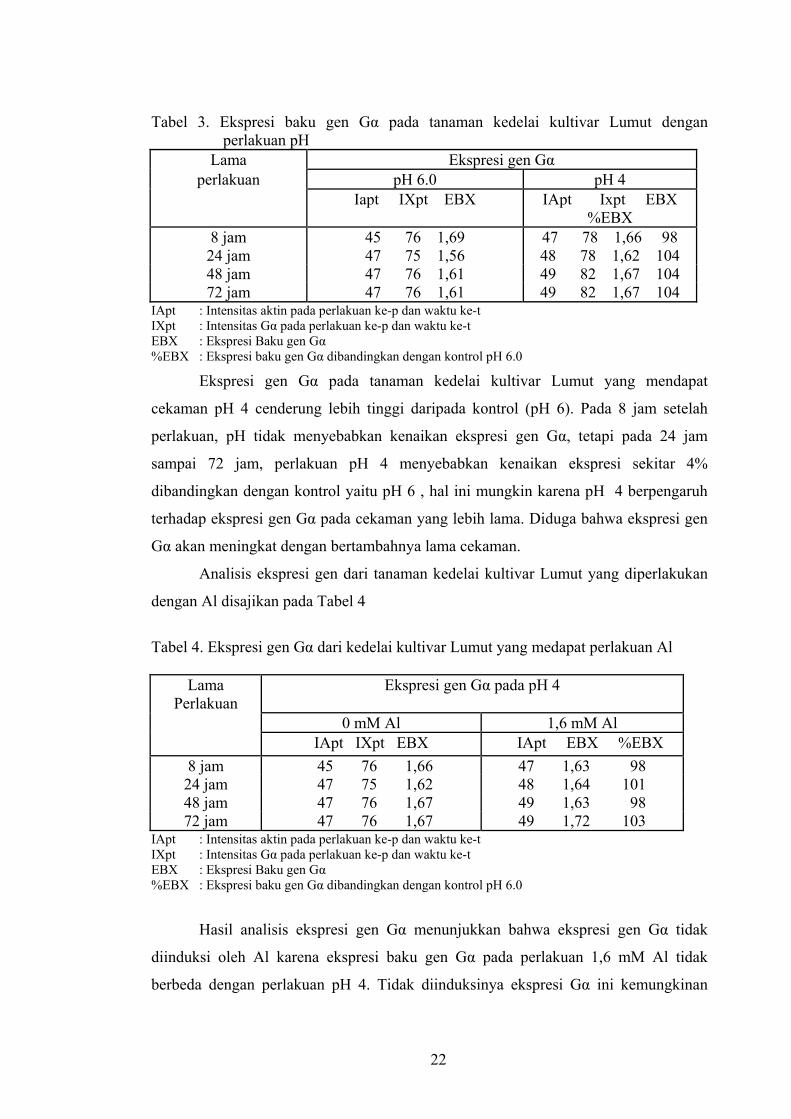
Tabel 3. Ekspresi baku gen Gα pada tanaman kedelai kultivar Lumut dengan perlakuan pH
Lama Ekspresi gen Gα perlakuan pH 6.0 pH 4
Iapt IXpt EBX IApt Ixpt EBX %EBX
8 jam 45 76 1,69 47 78 1,66 98 24 jam 47 75 1,56 48 78 1,62 104 48 jam 47 76 1,61 49 82 1,67 104 72 jam 47 76 1,61 49 82 1,67 104
IApt : Intensitas aktin pada perlakuan ke-p dan waktu ke-t IXpt : Intensitas Gα pada perlakuan ke-p dan waktu ke-t EBX : Ekspresi Baku gen Gα %EBX : Ekspresi baku gen Gα dibandingkan dengan kontrol pH 6.0
Ekspresi gen Gα pada tanaman kedelai kultivar Lumut yang mendapat
cekaman pH 4 cenderung lebih tinggi daripada kontrol (pH 6). Pada 8 jam setelah
perlakuan, pH tidak menyebabkan kenaikan ekspresi gen Gα, tetapi pada 24 jam
sampai 72 jam, perlakuan pH 4 menyebabkan kenaikan ekspresi sekitar 4%
dibandingkan dengan kontrol yaitu pH 6 , hal ini mungkin karena pH 4 berpengaruh
terhadap ekspresi gen Gα pada cekaman yang lebih lama. Diduga bahwa ekspresi gen
Gα akan meningkat dengan bertambahnya lama cekaman.
Analisis ekspresi gen dari tanaman kedelai kultivar Lumut yang diperlakukan
dengan Al disajikan pada Tabel 4
Tabel 4. Ekspresi gen Gα dari kedelai kultivar Lumut yang medapat perlakuan Al
Lama Perlakuan
Ekspresi gen Gα pada pH 4
0 mM Al 1,6 mM Al IApt IXpt EBX IApt EBX %EBX
8 jam 45 76 1,66 47 1,63 98 24 jam 47 75 1,62 48 1,64 101 48 jam 47 76 1,67 49 1,63 98 72 jam 47 76 1,67 49 1,72 103
IApt : Intensitas aktin pada perlakuan ke-p dan waktu ke-t IXpt : Intensitas Gα pada perlakuan ke-p dan waktu ke-t EBX : Ekspresi Baku gen Gα %EBX : Ekspresi baku gen Gα dibandingkan dengan kontrol pH 6.0
Hasil analisis ekspresi gen Gα menunjukkan bahwa ekspresi gen Gα tidak
diinduksi oleh Al karena ekspresi baku gen Gα pada perlakuan 1,6 mM Al tidak
berbeda dengan perlakuan pH 4. Tidak diinduksinya ekspresi Gα ini kemungkinan
22
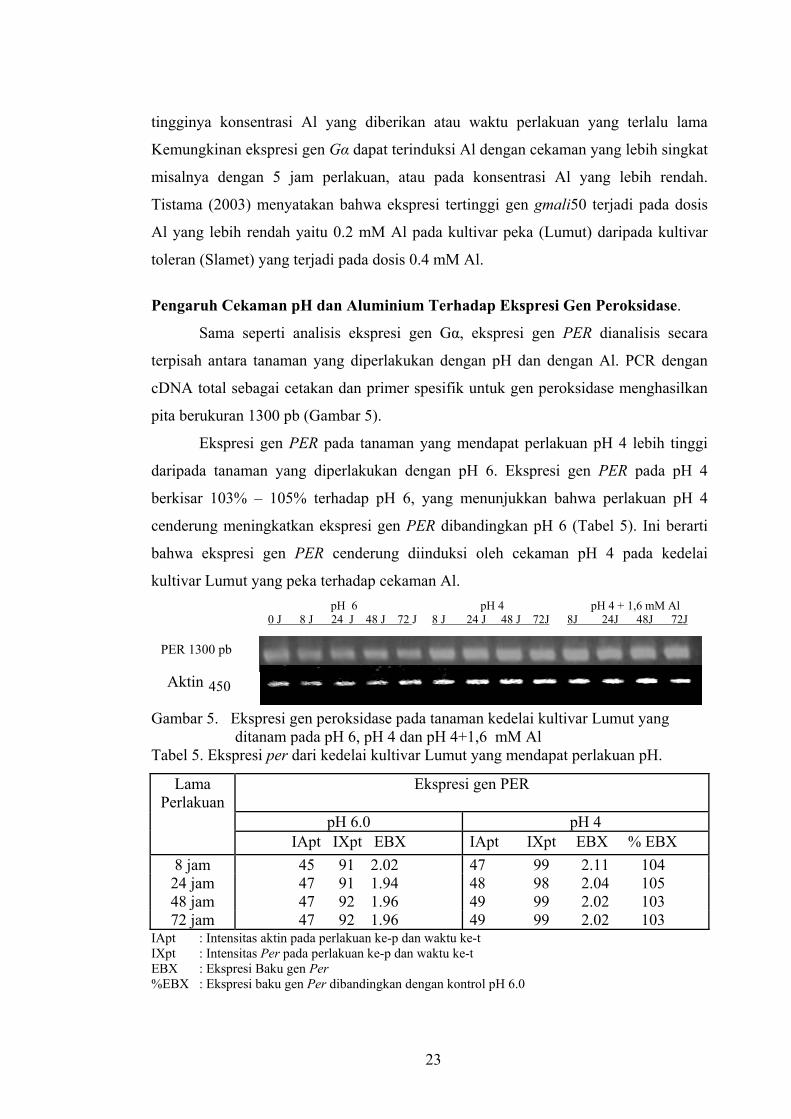
tingginya konsentrasi Al yang diberikan atau waktu perlakuan yang terlalu lama
Kemungkinan ekspresi gen Gα dapat terinduksi Al dengan cekaman yang lebih singkat
misalnya dengan 5 jam perlakuan, atau pada konsentrasi Al yang lebih rendah.
Tistama (2003) menyatakan bahwa ekspresi tertinggi gen gmali50 terjadi pada dosis
Al yang lebih rendah yaitu 0.2 mM Al pada kultivar peka (Lumut) daripada kultivar
toleran (Slamet) yang terjadi pada dosis 0.4 mM Al.
Pengaruh Cekaman pH dan Aluminium Terhadap Ekspresi Gen Peroksidase.
Sama seperti analisis ekspresi gen Gα, ekspresi gen PER dianalisis secara
terpisah antara tanaman yang diperlakukan dengan pH dan dengan Al. PCR dengan
cDNA total sebagai cetakan dan primer spesifik untuk gen peroksidase menghasilkan
pita berukuran 1300 pb (Gambar 5).
Ekspresi gen PER pada tanaman yang mendapat perlakuan pH 4 lebih tinggi
daripada tanaman yang diperlakukan dengan pH 6. Ekspresi gen PER pada pH 4
berkisar 103% – 105% terhadap pH 6, yang menunjukkan bahwa perlakuan pH 4
cenderung meningkatkan ekspresi gen PER dibandingkan pH 6 (Tabel 5). Ini berarti
bahwa ekspresi gen PER cenderung diinduksi oleh cekaman pH 4 pada kedelai
kultivar Lumut yang peka terhadap cekaman Al. pH 6 pH 4 pH 4 + 1,6 mM Al 0 J 8 J 24 J 48 J 72 J 8 J 24 J 48 J 72J 8J 24J 48J 72J
PER 1300 pb Aktin 450 Gambar 5. Ekspresi gen peroksidase pada tanaman kedelai kultivar Lumut yang
ditanam pada pH 6, pH 4 dan pH 4+1,6 mM Al Tabel 5. Ekspresi per dari kedelai kultivar Lumut yang mendapat perlakuan pH.
Lama Perlakuan
Ekspresi gen PER
pH 6.0 pH 4 IApt IXpt EBX IApt IXpt EBX % EBX
8 jam 45 91 2.02 47 99 2.11 104 24 jam 47 91 1.94 48 98 2.04 105 48 jam 47 92 1.96 49 99 2.02 103 72 jam 47 92 1.96 49 99 2.02 103
IApt : Intensitas aktin pada perlakuan ke-p dan waktu ke-t IXpt : Intensitas Per pada perlakuan ke-p dan waktu ke-t EBX : Ekspresi Baku gen Per %EBX : Ekspresi baku gen Per dibandingkan dengan kontrol pH 6.0
23

Perlakuan Al cenderung menyebabkan penghambatan ekspresi gen PER. Hal
ini dapat dilihat pada hasil analisis ekspresi gen yang disajikan pada Tabel 6. Pada
berbagai lama perlakuan, ekspresi gen PER dari tanaman yang mendapat perlakuan pH
4 + 1,6 mM Al lebih rendah atau sama dengan ekspresi gen tersebut pada tanaman
yang mendapat perlakuan pH 4. Sama seperti ekspresi gen Gα, terdapat
kecenderungan bahwa gen PER diinduksi oleh pH 4 tetapi tidak oleh Al. Cekaman pH
4 dan pH 4+1,6 mM Al juga tidak menginduksi ekspresi gen GST 12 pada kedelai
Lumut (Sawitri 2007).
Tabel 6. Ekspresi gen PER dari tanaman kedelai kultivar Lumut yang mendapat
perlakuan pH 4 dan Al Lama Perlakuan
Ekspresi gen PER pada pH 4 dan Al
0 mM Al 1,6 mM Al Iapt Ixpt EBX IApt IXpt EBX % EBX 8 jam 47 99 2.11 49 98 2.00 99.01 24 jam 48 98 2.04 50 99 1.98 102.06 48 jam 49 99 2.02 49 99 2.02 103.06 72 jam 49 99 2.02 50 99 1.98 101.02
IApt : Intensitas aktin pada perlakuan ke-p dan waktu ke-t IXpt : Intensitas Per pada perlakuan ke-p dan waktu ke-t EBX : Ekspresi Baku gen Per %EBX : Ekspresi baku gen Per dibandingkan dengan kontrol pH 6.0
24

SIMPULAN
Ekspresi gen Gα dan PER pada tanaman kedelai kultivar Lumut cenderung
diinduksi oleh cekaman pH 4 tetapi tidak oleh aluminium.
SARAN
Dilakukan penelitian lanjutan dengan konsentrasi Al yang lebih rendah dan
dengan mengurangi lama cekaman.
25

DAFTAR PUSTAKA
Aharon GS, Gelli A, Snedden WA, Blumwald E. 1998. Activation of a plant plasma
membrane Ca2+ channel by TGα 1, a heterotrimeric G protein α subunit homologue. FEBS lett 424: 17-21.
Anwar S, Suharsono, Jusuf M. 1999. The molecular respone of soybean roots to
aluminium stress. One day seminar on biotechnology. IPB. Bogor. April 14. Anwar S, Jusuf M, Suharsono, Sopandie D. 2000. Pengklonan gen yang diinduksi oleh
aluminium pada kedelai. J Bioteknol Indonesia 5 (1): 7-16. Beffa R et al. 1995. Cholera toxin elevates pathogen resistance and induces
pathogenesis-related gene expression in tobacco. EMBO J 23: 5753-5762. Bischoff F, Molendjik A, Rajendrakumar CSV, Plame K. 1999. GTP-binding protein
in plants. Cell Mol Life Sci 55: 233-256. Blokhina O, Virolainen E, Fagerstedt KV. 2002. Antioxidants damage and oxygen
deprivation stress: a review. Ann of Botany 91: 179-194. Cakmak I, Horst W J. 1991. Effect of aluminum peroxidation, superoxide dismutase,
catalase and peroxidase activity in root tips of soybean (Glycine max). Plant Physiol 83: 463-468.
Christiansen-Weniger C, Gronemon AF, Van Veen JA. 1992. Associative N2 fixation
and root exudation of organic acids from wheat cultivars of different aluminum tolerance. Plant Soil 139: 167-174.
Cote GG, Crain RC. 1993. Biochemistry of Phosphoinositides. Annu Rev Plant
Physiol 44: 333-356. Delhaize R, Ryan PR. 1995. Aluminum toxicity and tolerance in plants. Plant Physiol
107: 315-321. Dengenhardt J, Larsen PB, Howell SH, Kochian LV. 1998. Aluminum resistance in
the Arabidopsis mutant alr-104 is caused by an aluminum increase in rhizospHere pH. Plant Physiol. 122: 657-666.
Dunford HB. 1991. Horseradish peroxidase: structure and kinetic properties. In:
Everse J, Grisham MB, (ed). Peroxidases in Chemistry and Biology. CRC Press. Boca Raton, Florida. : 1-24.
Ezaki B, Gardner RC, Matsumoto H. 2000. Expression of aluminum-induced genes in transgenic Arabidopsis plants can ameliorate aluminum stress and/or oxidative stress. Plant Physiol 122: 657-666.
26

Fairley-Grenot K, Assman SM. 1991. Evidence for G protein regulation of inward potassium ion channel current in guard cells of Fava bean. Plant Cell 3: 1037-1044.
Fujisawa Y, Kato H, Iwasaki Y. 2001. Structure and function of heterotrimeric G
protein in plants. Plant Cell Physiol 42 (8): 789-794. Galvez L, Clark RB. 1991. Nitrate and ammonium uptake changes for Al-tolerant and
Al-sensitive sorghum (Sorghum bicolor) genotypes grown with and without aluminum. Plant Soil 134: 179-188.
Hamm HE. 1998. The many faces of G protein signaling. J Biol Chem 273(2): 669-
672. Haug, A. 1984. Molecular aspects of aluminum toxicity. CRC Crit Rev Plant Sci 1:
345-373. Helyar KR. 1998. The symptoms and effect on plants of nutrient disorders in acid
soils. In: 25 years of the Riverina Outlook Conference, Wagga Wagga,1973-1998 (online). http://www.regional.org.au/au/roc/1981/roc198147.htm. Tanggal 1 Februari 2006
Hiraga S, Sasaki K, Ito H, Ohashi Y, Matsui H. 2001. A large family pf class III plant
peroxidases. Plant Cell Physiol 42: 462-468. Ismail I G, Effendi S. 1993. Pertanaman Kedelai pada Lahan Kering. Kedelai. Di
dalam: Somaatmaja S, Ismunadji M, Sumarno, syam M, Manurung SO, Yuswadi, editor. Kedelai. Bogor: Balai Penelitian dan Pengembangan Pertanian. Pusat Penelitian dan Pengembangan Tanaman Pangan. hlm 103-119.
Iwasaki Y, Kato T, Kaidoh T, Ishikawa A, Asahi T. 1997. Characterization of the
putative α subunit of a heterotrimeric G protein in rice. Plant Mol Biol 34: 563-572.
Johnson JP, Cerver BF, Baligar VC. 1997. Expression of aluminum tolerance
transferred from Atlas 66 to hard winter wheat. Crop Sci 37: 103-108. Jones DL, Kochian LV. 1995. Aluminum inhibition of the inositol 1,4,5-triphosphate
signal transduction pathway in wheat roots: A role in aluminum toxicity. Plant Cell 7: 1913-1922.
Jusuf M, Suharsono, Sopandie D. 1999. Molecular biology of Soybean tolerance to
aluminum stress. Report of Graduate Team Research Grant, URGE Project, Batch II: Directorate General of Higher Education. Jakarta
Kennedy IR. 1992. Acid Soil and Acid Rain. Second Edition. John Wiley & Sons Inc. New York. 254.
27

Kinraide TB. 1991. Identity of the rhizotoxic aluminum species. Plant Soil 134: 167-
178. Kochian LV. 1995. Cellular mechanisms of aluminum toxicity and resistance in
plants. Annu Rev Plant Physiol Plant Mol Biol 46: 237-260.
Krauss 2001. Inositol triphoshate and calsium signaling. Nature 251: 315-325. Legendre L, Heinstein PF, Low PS. 1992. Evidence for participation of GTP-binding
protein in elicitation of the rapid oxidative burst in cultured soybean cells. J Biol Chem 267: 20140-20147.
Legendre L, Henstein PF, Low PS. 1993. Phospholipase C activation during elicitation
of the oxcidative burst in cultured plant cell. J Biol Chem 268: 24559-24563. Li W, Assmann SM. 1993. Characterization of a G protein-regulated outward
potassium current in mesophyl cell Vicia faba. Proc Natl Acad Sci USA 90: 262-266.
Ma H. 1994. GTP-binding proteins in plants: a new members of an old family. Plant
Mol Biol 26: 1611-1636. Ma JK, Zheng SJ, Matsumoto H. 1997. Spesific secretion of citric acid induced by Al
stress in Cassia tora L. Plant Physiol 38: 1019-1025. Mahady GB, Liu C, Beecher CWW. 1998. Involvement of protein kinase C and G
protein in the signal transduction of benzo phenanthridine alkaloid biosynthesis. Phytochem 48: 93-102.
Marschner H. 1986. Mineral Nutrition of Higher Plant. Academic Press Harcourt
Brace and Company. London. Martin RB. 1986. The chemistry of aluminum as related to biology and medicine.
Clin Chem 32: 1797-1806. Martin RB. 1988. Bioinorganic chemistry of aluminum. 1-57 Di dalam: Sigel H, Sigel
A, editors. Metal Ions in Biological Systems: Aluminium and Its Role in Biology 24. New York:Marcel-Dekker.
Mashuda 2006. Ekspresi Gen Gα dan GST pada Kedelai Kultivar Slamet yang
Mendapat Cekaman Aluminium [Tesis]. Sekolah Pascasarjana Institut Pertanian Bogor.
Matsumoto H, Hirasawa E, Morimura S, Takahashi E. 1976. Localization of aluminum in tea leaves. Plant Cell Physiol 17: 627-631.
28

Matsumoto H. 1991. Biochemical mechanism of the toxicity of aluminium and the sequestration of aluminium in plant cell. Plant Cell Physiol 21: 825-838.
Matsumoto H. 2000. Cell biology of aluminum toxicity and tolerance in higher plants.
Internat Rev Cyt 200: 1-46. Mc Laughin S, Carolyn B, Gennady D, Michel G. 1996. The importance of lipid-
protein interaction in signal transduction through the calcium-phospholipid second messenger system. NATO ASI Series H Cell Biol 95.
Miyasaka SC, Buta JG, Hawell RK, Foy CD. 1991. Mechanism of aluminum tolerance
in snapbean: root exudation of citric acid. Plant Physiol 96: 737-743. Moller T, Bailar JL, Kleinberg J, Guss CO, Castellion ME and Molz C. 1984.
Chemistry with Inorganik Qualitative Analysis Acid Press, Inc Orlando. Notohadiprawiro T. 1983. Persoalan Tanah Masam dalam Pembangunan Pertanian
Indonesia. Bull Faperta 18: 44-47. Osaki M, Watanabe T, Tadano T. 1997. Beneficial effect of aluminum growth of plant
adapted to low pH soil. Soil Sci Plant Nutr 43 (3): 551-563. ∅stergaard L, Teilum K, Mirza O, Mattsson O, Petersen M, Welinde KG, Mundy J,
Gajhede M, Henriksen A. 2000. Arabidopsis ATP A2 peroxidase. Expression and high-resolution structure of a plant peroxidase with implications for lignification. Plant Mol Biol 44: 231-243.
Pellet DM, Grunes DL, Kochian LV. 1995. Organic acid exudation as an aluminum-
tolerance mechanism in maize (Zea mays L.). Planta 196: 788-795. Peskan T, Oelmuller R. 2000. Heterotrimeric G-protein β – subunit is localized in the
plasma membrane and nuclei tobacco leaves. Plant Mol Biol 42: 915-922. Pineros MA, Jurandir V, Magalhaes, Vera M, Alves C, Kochian LV. 2002. The
physiology and biophysics of an aluminum tolerance mechanism based on root citrate exudation in maize. Plant Physiol 129: 1194-1206.
Prihardi DP, ER Shipe, SU Wallace. 1995. Screening selected soybean genotype for
aluminum tolerance. Indon J Trop Agric 6(1): 11-12. Richards KD, Schott EJ, Sharma YK, Davis KR, Gardner RC. 1998. Aluminum
induces oxidative stress genes in Arabidopsis thaliana. Plant Physiol 116: 409-418.
Ryan PR, JM Ditomaso, Kochian LV. 1993. Aluminum toxicity in roots: an
investigation of spatial sensitivity and the role of the root cap. J Exp Bot 44 (259): 437-446.
29

Ryan PR, Kinraide TB, Kochian LV. 1994. Al3+Ca2+ interaction in aluminum rhizotoxicity inhibition of root growth is not caused by reduction of calsium uptake. Planta 192: 98-103.
Ryan PR, Delhaize E, Randall PJ. 1995. Malate efflux from root apices: evidence for a
general mechanism of Al-tolerance in wheat. Aust J Plant Physiol 22: 531-536. Sasaki M, Kasai M, Yamamoto Y, Matsumoto H. 1992. Root elongation and ion flux
of wheat varieties differing in aluminum tolerance. Plant Cell Wall as Biopolimers with Physiological Function. 401-403.
Sasaki M, Kasai M, Yamamoto Y, Matsumoto H. 1994. Comparation of the early
response to aluminum stress between tolerant and sensitive wheat cultivars: Root growth, aluminum content and efflux of K+. J Plant Nutr 17(7): 1275-1288.
Sawitri SM 2007. Ekspresi gen Gα dan GST pada kedelai kultivar Lumut yang
mendapat cekaman aluminium. [Tesis]. Bogor: Sekolah Pascasarjana Institut Pertanian Bogor.
Silva IR, Smyth TJ, Rarer CD, Carter TE, Rutty T. 2001. Differential aluminum
tolerance in soybean: an evaluation of the role of organic acid. Plant Physiol 112: 200-210.
Sopandie D, Jusuf M, Hamim, Supiatno. 1996. Fisiologi dan Genetik Daya Adaptasi
Kedelai terhadap Cekaman Kekeringan dan pH Rendah dengan Al tinggi. Laporan Riset Unggulan Terpadu (RUT I).
Suharsono 2002. Konstruksi pustaka genome kedelai kultivar Slamet. Hayati 9(3): 67-
70. Suharsono 2007. Pembuatan pustaka genom kedelai kultivar Lumut di dalam fage
lambda Biosfera (inpress) Suharsono U et al. 2002. The heterotrimeric G protein α subunit act upstream of the
small GTPase Rac in disease resistance of rice. Proc Natl Acad Sci 99 (20): 13307-13312.
Suharsono, Jusuf M, Anwar S, Widyastuti U. 2003. Isolasi dan Karakterisasi Gen-Gen
dari Tanaman Kedelai yang Mendapat Cekaman Aluminium. Laporan Riset Unggulan Terpadu VIII Bidang Bioteknologi Jakarta. Kementrian Riset dan Teknologi dan Lembaga Ilmu Pengetahuan Alam.
Suharsono, Jusuf M, Paserang AP. 2006. Analisis ragam heritabilitas dan pendugaan
kemajuan seleksi populasi F2 dari persilangan kedelai kultivar Slamet X Nokonsawon. J Tan Tropika 9(2): 86-93.
30

Suharsono, Jusuf M, Dasumiati. 2007. Analisis ragam dan seleksi populasi F3 dari persilangan kedelai kultivar Slamet X Nokonsawon. J Tan Tropika 10(1): 21-28.
Syarifuddin A, Abdurahman A. 1993. dalam Anwar 1999. Optimasi pemanfaatan
sumberdaya lahan berwawasan lingkungan. Prosiding Simposium Penelitian Tanaman Pangan III. Pusat Penelitian dan Pengembangan Tanaman Pangan dan Badan Litbang DEPTAN. Jakarta/Bogor 23-25 Agustus 1993.
Tang Y, Garvin DF, Kochian LV, Sorrells ME, Carver BF. 2002. Physiological
genetics of aluminum tolerance in the wheat cultivar atlas 66. Crop Sci 42: 1541-1546.
Taylor GJ. 1991. Current views of the aluminum stress response:The physiological basis of tolerance.Curr.Top Plant Biochem Physiol 10: 57-93.
Tistama R. 2003. Transkripsi gmali14 dan gmali50 pada Kedelai [Glycine max (L)
Merryl] yang Mendapat Cekaman Beberapa Logam dan Garam. [Tesis]. Bogor: Sekolah Pascasarjana Institut Pertanian Bogor.
Yamamoto Y, Ono K, Mamatsuka K, Kasai M, Matsumoto H. 1992. Growth inhibiton
by aluminum is alleviated by phosphate starvation in cultured tobacco cells. Plant cell walls as biopolymers with physiological function. Yamada Science Foundation. Osaka japan.
Yuniati R 2000. Pengklonan cDNA tanaman kedelai (Glycine max (L.) Merryl)
varietas Slamet yang diinduksi cekaman aluminium (tesis) Bogor. Institut Pertanian Bogor.
Van HL, Kuraishi S, Sakurai N. 1994. Aluminum-induced rapid root inhibition and changes in cell-wall components of squash seedling. Plant Physiol 106: 971-976.
Van Wambeke A. 1976. Formation, distributon and consequence of acid soil of agricultured development. in Wright MJ and SA Ferari, eds Plants adaptation to mineral stress in problem soils. Spec Publ Cornell Univ Agric Exp Stn Ithaca, New York. 15-24.
Weiss CA, White E, Huang H, Ma H. 1997. The Gα protein sub unit (G P α 1) is
associated with the endoplasmic reticulum and the plasma membrane in meristematic cells of Arabidopsis and Cauliflower. FEBS Lett 407: 361-367.
Wenzl P, Platino GM, Chaves AL, Meyer JE, Rao IM. 2001. The high level of
aluminum resistance in signalgrass is not associated with known mechanism of aluminum detoxification in root apices. Plant Physiol 125: 1473-1484.
Widjaja-Adhi, I P G. 1985. Pengapuran tanah masam untuk kedelai, p. 171-188.
Dalam S. Somaatmadja (ed). Kedelai. Pusat Penelitian dan Pengembangan Pertanian. Bogor.
31
32
Lampiran 1 Data Baku Pertambahan Panjang akar
pH 6 pH 4+1.2 mM Al 8jam 24 jam 48 jam 72 jam 8 jam 24 jam 48 jam 72 jam 0.4 1.3 2.1 2.9 0.00 0.10 0.20 0.30 0.4 1.3 2.1 2.9 0.10 0.10 0.20 0.30 0.20 1.40 2.10 2.80 0.00 0.30 0.40 0.40 ulangan 1 0.40 1.30 1.90 2.50 ulangan 1 0.00 0.30 0.30 0.40 0.50 1.20 1.90 2.70 0.10 0.20 0.30 0.30 0.30 1.40 2.00 2.80 0.10 0.20 0.20 0.30 0.40 1.30 2.10 2.90 0.00 0.30 0.30 0.30 0.50 1.40 1.9 2.70 0.10 0.10 0.20 0.30 0.60 1.10 1.9 2.80 0.10 0.20 0.20 0.30 0.40 1.30 2 3 0.10 0.20 0.20 0.30 rata-rata 0.41 1.3 2 2.8 rata-rata 0.06 0.20 0.25 0.32 0.5 1.20 1.7 2.50 ulangan 2 0.10 0.20 0.20 0.30 0.5 1.20 1.9 2.90 0.10 0.20 0.30 0.30 0.30 1.10 1.90 2.80 0.00 0.10 0.20 0.30 0.40 1.30 2.20 2.90 0.00 0.10 0.20 0.30 ulangan 2 0.50 1.30 2.00 3.30 0.00 0.10 0.20 0.30 0.60 1.20 1.90 2.90 0.00 0.10 0.20 0.30 0.40 1 2.10 3.10 0.00 0.10 0.20 0.30 0.60 1.3 1.90 2.70 0.10 0.20 0.30 0.30 0.50 1.2 2.30 2.90 0.10 0.20 0.30 0.30 0.40 1.3 1.90 3.00 0.00 0.10 0.20 0.20 rata-rata 0.47 1.21 1.98 2.90 rata-rata 0.04 0.14 0.23 0.29 pH 4 pH 4+1.6 mM Al 8 jam 24 jam 48 jam 72 jam 8 jam 24 jam 48 jam 72 jam 0.40 0.70 1.20 1.90 0.00 0.1 0.1 0.2 0.20 0.60 1.00 1.60 0.10 0.1 0.2 0.2 0.10 0.50 0.90 1.50 0.00 0 0.1 0.2 0.30 0.70 1.20 1.60 0.00 0.1 0.2 0.2 ulangan 1 0.20 0.60 1.20 1.40 ulangan 1 0.10 0.1 0.2 0.3 0.40 0.90 1.20 1.6 0.10 0.10 0.20 0.20 0.30 0.70 1.10 1.60 0.00 0.10 0.20 0.20 0.10 0.50 1.00 1.50 0.10 0.10 0.20 0.20 0.20 0.60 1.20 1.50 0.00 0.00 0.10 0.20 0.10 0.50 1.00 1.50 0.00 0.00 0.10 0.20 rata-rata 0.23 0.63 1.10 1.57 rata-rata 0.04 0.07 0.16 0.21 0.30 0.60 1.20 1.40 0.10 0.10 0.20 0.20 0.30 0.70 1.30 1.40 0.10 0.10 0.20 0.20 0.20 0.50 0.90 1.50 0.00 0.00 0.10 0.20 0.30 0.60 0.90 1.30 0.00 0.10 0.10 0.20 ulangan 2 0.40 0.90 1.10 1.50 0.00 0.10 0.10 0.10 0.10 0.50 0.90 1.30 ulangan 2 0.00 0.10 0.10 0.20 0.20 0.60 1.10 1.50 0.10 0.10 0.20 0.20 0.20 0.70 1.00 1.40 0.10 0.10 0.20 0.20 0.30 0.70 1.20 1.90 0.10 0.10 0.20 0.20 0.20 0.70 1.10 1.50 0.10 0.10 0.20 0.20 rata-rata 0.25 0.65 1.07 1.47 rata-rata 0.06 0.09 0.16 0.19
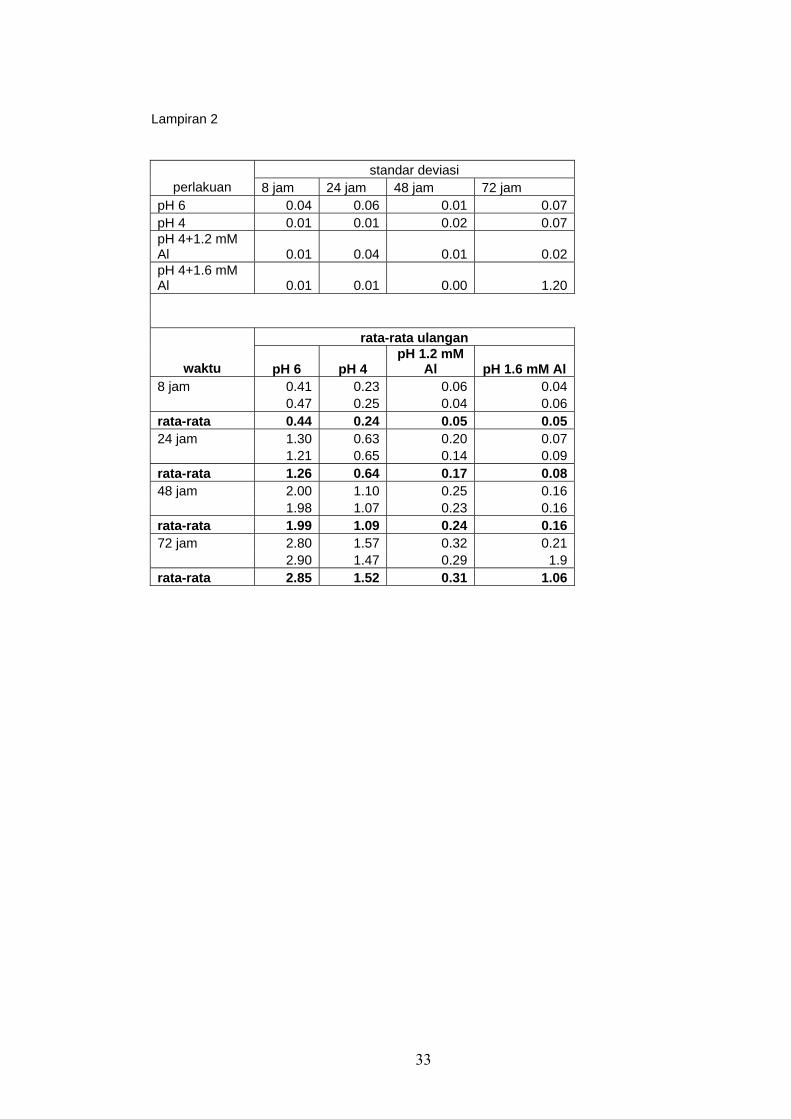
Lampiran 2
standar deviasi perlakuan 8 jam 24 jam 48 jam 72 jam
pH 6 0.04 0.06 0.01 0.07 pH 4 0.01 0.01 0.02 0.07 pH 4+1.2 mM Al 0.01 0.04 0.01 0.02 pH 4+1.6 mM Al 0.01 0.01 0.00 1.20
rata-rata ulangan
waktu pH 6 pH 4 pH 1.2 mM
Al pH 1.6 mM Al 8 jam 0.41 0.23 0.06 0.04 0.47 0.25 0.04 0.06 rata-rata 0.44 0.24 0.05 0.05 24 jam 1.30 0.63 0.20 0.07 1.21 0.65 0.14 0.09 rata-rata 1.26 0.64 0.17 0.08 48 jam 2.00 1.10 0.25 0.16 1.98 1.07 0.23 0.16 rata-rata 1.99 1.09 0.24 0.16 72 jam 2.80 1.57 0.32 0.21 2.90 1.47 0.29 1.9 rata-rata 2.85 1.52 0.31 1.06
33

0 Jam
Lampiran 3.
Foto tanaman kedelai kultivar Lum
ut pada berbagai perlakuan
8 Jam
24 Jam
48 Jam
72 Jam
34
Related Documents











